Local sensory processing of light determines basic features of shape and movement.
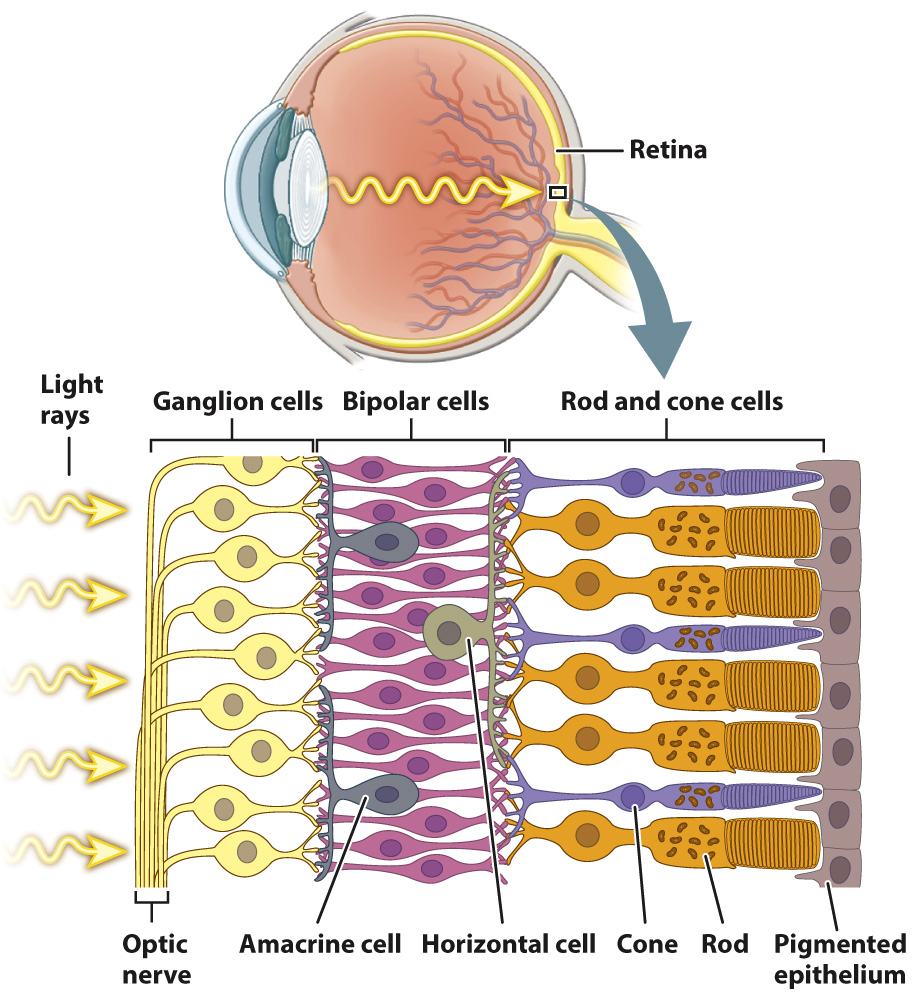
The rods and cones detect the intensity, color, and pattern of light entering the eye through the lens. However, it is their interaction with a highly ordered array of neurons in the retina that begins the first steps of visual sensory processing (Fig. 36.19). The retina consists of five layers of cells that form a signal-
776
777
The bipolar cells, in turn, synapse on to ganglion cells located on the front of the retina. If activated, ganglion cells transmit action potentials by the optic nerve (a cranial nerve) to the visual cortex in the brain, the part of the brain that processes images. The optic nerve begins at the front of the retina and exits at the back, creating an area without light-
In addition to these three primary layers of neurons, two additional sets of nerve cells combine information laterally across the retina. Horizontal cells communicate between neighboring groups of photoreceptors and bipolar cells, enhancing contrast and adjusting photoreceptor sensitivity to light levels. Although their function is less well understood, we know that amacrine cells communicate between neighboring bipolar cells and ganglion cells, enhancing motion detection and likely reinforcing the adjustment of photoreceptor light sensitivity by horizontal cells.
Photoreceptor cells, bipolar cells, ganglion cells, horizontal cells, and amacrine cells form a neural network that processes visual information before it is sent to the brain for further processing and interpretation. Visual signals from approximately 131 million photoreceptors converge to approximately 1 million ganglion cell neurons that communicate with the brain. Properties of this network were first studied in the mammalian retina by the American neurophysiologists Stephen Kuffler, David Hubel, and Torsten Wiesel in the 1950s and 1960s (Fig. 36.20). Hubel and Wiesel shared the 1981 Nobel Prize in Physiology or Medicine for their later work on how visual information is processed in the brain.
HOW DO WE KNOW?
FIG. 36.20
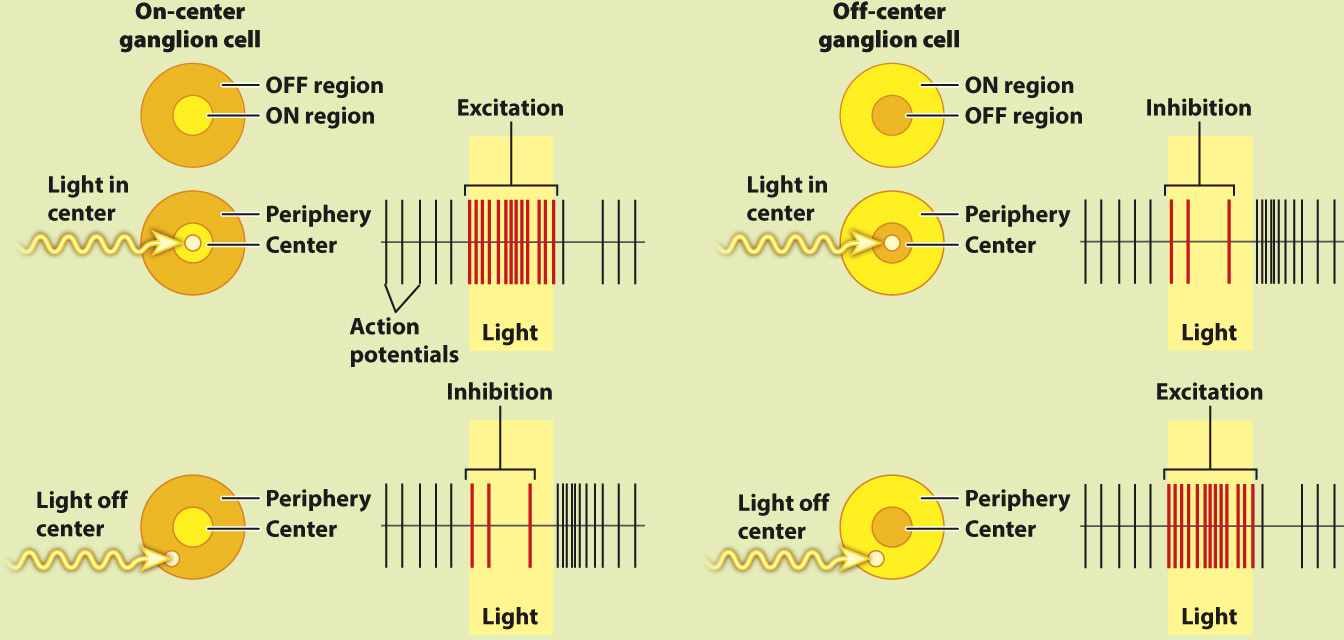
How does the retina process visual information?
BACKGROUND In the 1950s, the American neurophysiologist Stephen Kuffler was interested in understanding how the retina helps to process light information before it is sent to the brain. He focused on the activity of ganglion cells in the retina because they receive input from the photoreceptors and bipolar cells.
EXPERIMENT Kuffler stimulated different regions of a cat’s retina with localized points of light while recording the action potentials produced by ganglion cells.
RESULTS Kuffler found that there are two types of ganglion cell: on-
FOLLOW-
SOURCES Kuffler, S. W. 1953. “Discharge Patterns and Functional Organization of Mammalian Retina.” Journal of Neurophysiology 16:37–