The first trimester includes cleavage, gastrulation, and organogenesis.
How does a single cell become a complex, multicellular organism? Development depends on a precisely orchestrated set of cell divisions, movements, and specializations coordinated by both maternal and zygotic genetic instructions (Chapter 20). In this section, we focus on three key processes in early embryonic development: cleavage, gastrulation, and organogenesis.
After fertilization, the single-
914
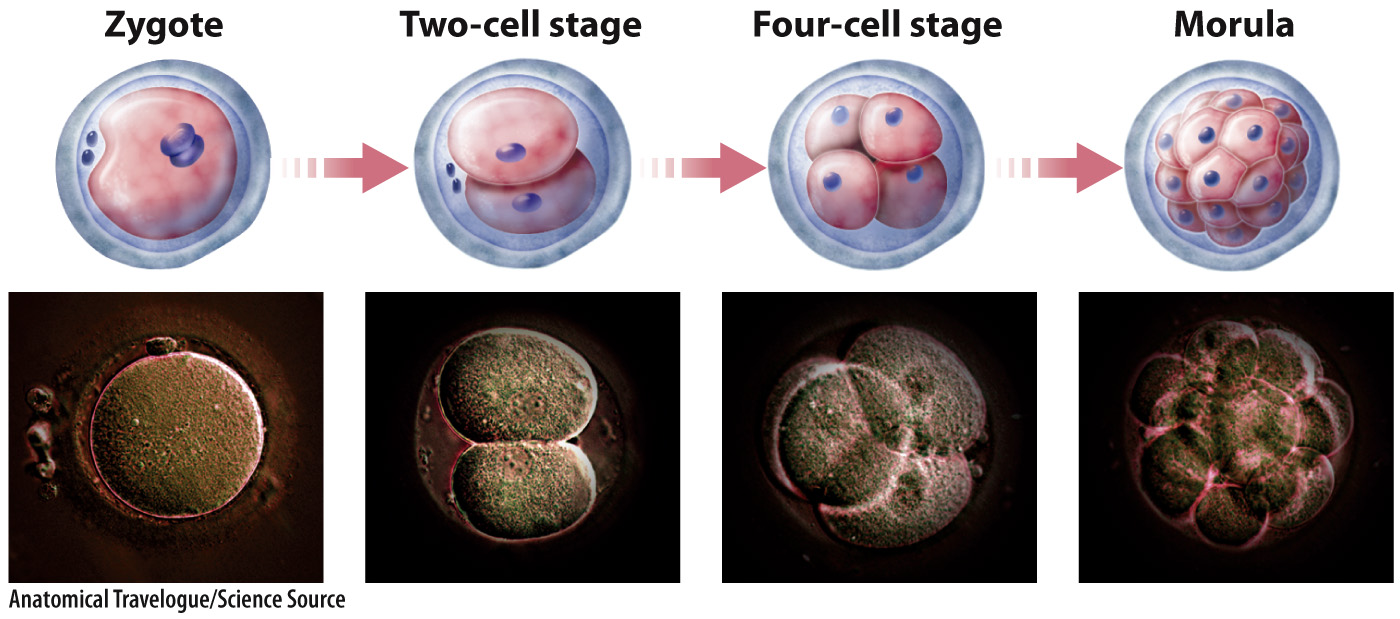
Cleavage in mammals is unique in several respects. It is slow compared with cleavage in other organisms. In addition, in mammals the early divisions do not occur at the same time, so it is not unusual to find an odd number of cells in the developing embryo. Finally, zygotic transcription begins very early, as early as the first mitotic division, or two-
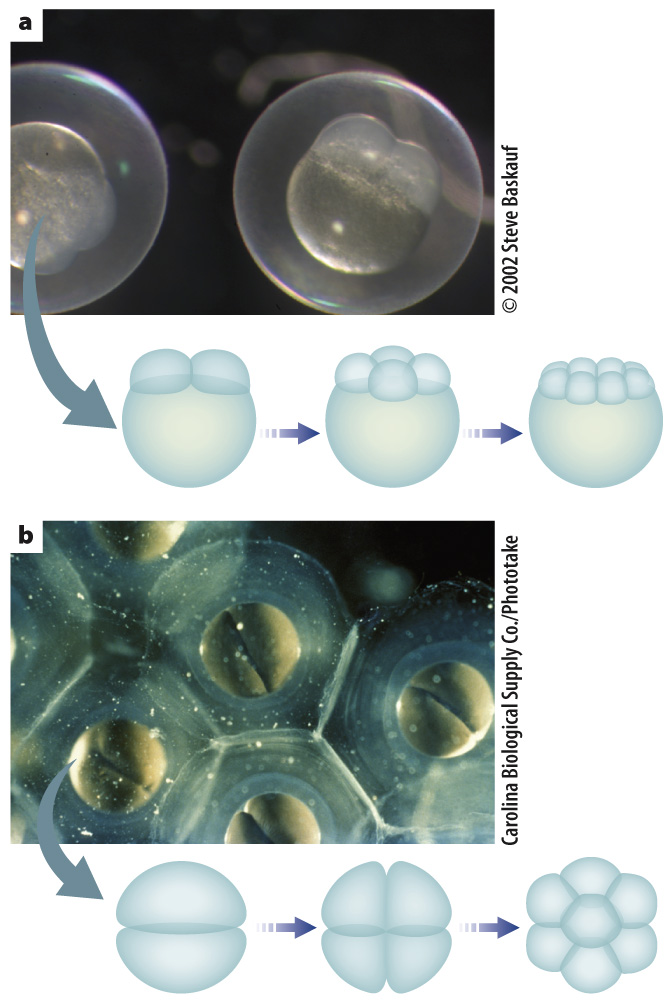
The pattern of cleavage is different for different organisms. It is determined in part by the amount of yolk present in the egg because the presence of yolk inhibits cell division (Fig. 42.22). In animals with lots of yolk, such as insects, fish, and birds, cell division does not extend through the entire egg (Fig. 42.22a). In animals with very little yolk, such as mammals and sea urchins, cell division extends completely through the egg (Fig. 42.22b). In the absence of sufficient yolk, the developing embryo needs another source of nutrition, which in humans and other placental mammals is provided by the placenta.
Cleavage results in a solid ball of cells called a morula (see Fig. 42.21). Further cell divisions result in a fluid-
Implantation marks the beginning of pregnancy, the carrying of one or more embryos in the uterus. It lasts approximately 38 weeks from fertilization. Because the time of fertilization is often not known, human pregnancy is typically measured not from implantation but as 40 weeks from the first day of the last menstrual period. Pregnancy is divided into three periods called trimesters, each lasting about 3 months. The divisions between the three trimesters are arbitrary, but they provide a convenient way to follow the major changes that occur during pregnancy.
The first trimester is characterized by spectacular changes in the developing embryo under the influence of genes and developmental pathways that are largely conserved across vertebrates (Chapter 20). During this time, the embryo develops all its major organs and begins to be recognizably human. Toward the end of the first trimester, a human embryo is known as a fetus.
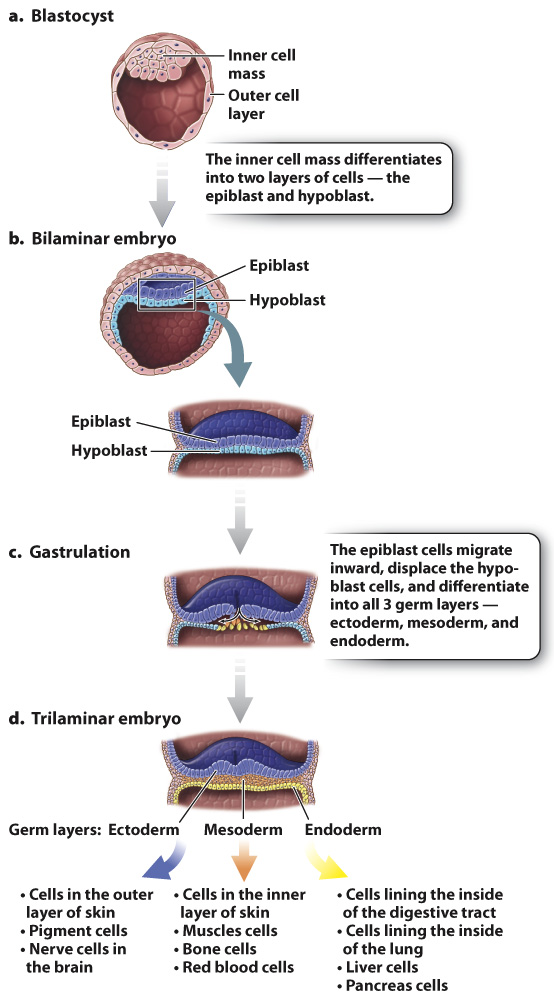
In mammals, such as humans, the blastocyst has an inner cell mass and an outer layer of cells (Fig. 42.23a). The inner cell mass becomes the embryo itself, and the outer cell layer forms part of the placenta. The placenta is composed of both maternal and embryonic cells and is the site of gas exchange, nutrient uptake, and waste excretion between maternal and fetal blood. The placenta also secretes the hormones estrogen and progesterone, which maintain the uterine lining during pregnancy. Meanwhile, the inner cell mass differentiates into a two-
Gastrulation is the second key developmental process in embryogenesis. It is a highly coordinated set of cell movements that leads to a fundamental reorganization of the embryo. In humans, cells of the epiblast divide and migrate inward (Fig. 42.23c). These migrating epiblast cells form three layers or sheets of cells called germ layers, which include the outer ectoderm, intermediate mesoderm, and inner endoderm (Fig. 42.23d). All of the adult tissues and organs are made from the three germ layers. At this stage, the embryo is known as a trilaminar (three-
As with cleavage, the details of gastrulation vary in different organisms. One notable difference is the fate of the opening through which cells migrate during gastrulation. In protostomes (“mouth first”), such as worms, mollusks, and arthropods, it becomes the mouth, whereas in deuterostomes (“mouth second”), such as vertebrates and sea urchins, it becomes the anus and the mouth forms later (Chapter 44).
Organogenesis, the third key developmental process, is the transformation of the three germ layers into all the organ systems of the body. The ectoderm becomes the outer layer of the skin and nervous system, the mesoderm makes up the circulatory system, muscle, and bone, and the endoderm becomes the lining of the digestive tract and lungs (Fig. 42.23d). Because all the basic structures and organ systems develop during the first trimester of pregnancy, it is during this time that the developing fetus is particularly vulnerable to toxins, drugs, and infections.
It is also during the first trimester that the embryo begins to develop male or female characteristics. In humans, sex determination is achieved by the presence or absence of the Y chromosome. The Y chromosome contains a gene, SRY, which encodes a protein called testis-
915