23.1 Biological Constraints on Conditioning
23-
Ever since Charles Darwin, scientists have assumed that all animals share a common evolutionary history and thus share commonalities in their makeup and functioning. Ivan Pavlov and John D. Watson, for example, believed the basic laws of learning were essentially similar in all animals. So it should make little difference whether one studied pigeons or people. Moreover, it seemed that any natural response could be conditioned to any neutral stimulus.
301
Limits on Classical Conditioning
In 1956, learning researcher Gregory Kimble proclaimed, “Just about any activity of which the organism is capable can be conditioned and … these responses can be conditioned to any stimulus that the organism can perceive” (p. 195). Twenty-
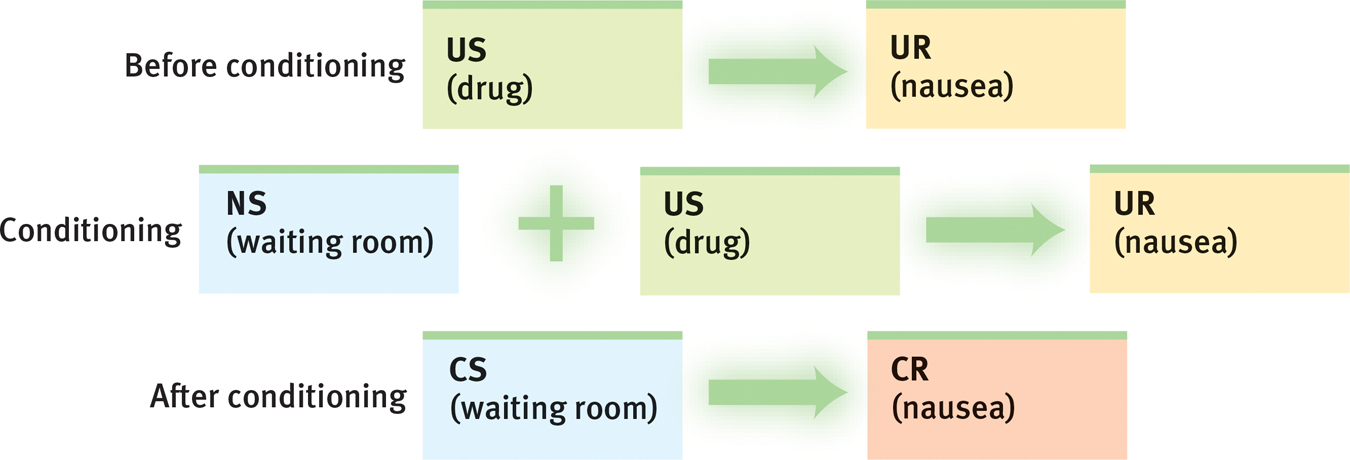
John Garcia was among those who challenged the prevailing idea that all associations can be learned equally well. While researching the effects of radiation on laboratory animals, Garcia and Robert Koelling (1966) noticed that rats began to avoid drinking water from the plastic bottles in radiation chambers. Could classical conditioning be the culprit? Might the rats have linked the plastic-

To test their hunch, Garcia and Koelling exposed the rats to a particular taste, sight, or sound (CS) and later also to radiation or drugs (US) that led to nausea and vomiting (UR). Two startling findings emerged: First, even if sickened as late as several hours after tasting a particular novel flavor, the rats thereafter avoided that flavor. This appeared to violate the Pavlovian notion that for conditioning to occur, the US must immediately follow the CS.

Second, the sickened rats developed aversions to tastes but not to sights or sounds. This contradicted the behaviorists’ idea that any perceivable stimulus could serve as a CS. But it made adaptive sense. For rats, the easiest way to identify tainted food is to taste it; if sickened after sampling a new food, they thereafter avoid it. This response, called taste aversion, makes it difficult to eradicate a population of “bait-
Humans, too, seem biologically prepared to learn some associations rather than others. If you become violently ill four hours after eating contaminated mussels, you will probably develop an aversion to the taste of mussels but usually not to the sight of the associated restaurant, its plates, the people you were with, or the music you heard there. (In contrast, birds, which hunt by sight, appear biologically primed to develop aversions to the sight of tainted food [Nicolaus et al., 1983].)
302
Garcia’s early findings on taste aversion were met with an onslaught of criticism. As the German philosopher Arthur Schopenhauer (1788–
“All animals are on a voyage through time, navigating toward futures that promote their survival and away from futures that threaten it. Pleasure and pain are the stars by which they steer.”
Psychologists Daniel T. Gilbert and Timothy D. Wilson, “Prospection: Experiencing the Future,” 2007
It is also a good example of experiments that begin with the discomfort of some laboratory animals and end by enhancing the welfare of many others. In one conditioned taste-
Such research supports Darwin’s principle that natural selection favors traits that aid survival. Our ancestors who readily learned taste aversions were unlikely to eat the same toxic food again and were more likely to survive and leave descendants. Nausea, like anxiety, pain, and other bad feelings, serves a good purpose. Like a low-

In an example illustrating how a CS can signal an important biological event, Japanese quail were conditioned to get sexually excited by a red light that signaled a receptive female’s arrival. Michael Domjan and his colleagues (2004) report that such conditioning is even speedier, stronger, and more durable when the CS is ecologically relevant—something similar to stimuli associated with sexual activity in the natural environment, such as the stuffed head of a female quail. In the real world, observes Domjan (2005), conditioned stimuli have a natural association with the unconditioned stimuli they predict.
The tendency to learn behaviors favored by natural selection may help explain why we humans seem naturally disposed to learn associations between the color red and sexuality. Female primates display red when nearing ovulation. In human females, enhanced bloodflow produces the red blush of flirtation and sexual excitation. Does the frequent pairing of red and sex—
- men tipped waitresses in red more generously than waitresses in any other color, and on a dating website, men were most likely to contact a woman wearing red (Guéguen & Jacob, 2013, 2014).
- women became more likely to select a red rather than a green shirt to wear when expecting to converse with an attractive rather than an unattractive man (Elliot et al., 2013).
- when at peak fertility, women are more likely to wear red or pink (Beall & Tracy, 2013). The red effect enhances men’s attraction only to younger, fertile (not menopausal) women (Schwarz & Singer, 2013).
- and it’s not just men: Women tend to perceive men as more attractive when seen in photos on a red background or in red clothing (Elliot et al., 2010; Roberts et al., 2010).

Romantic red In a series of experiments that controlled for other factors (such as the brightness of the image), men found women more attractive and sexually desirable when framed in red (Elliot & Niesta, 2008). The phenomenon has been found not only in North America and Europe, but also in the West African nation of Burkina Faso (Elliot et al., 2013).
303
A genetic predisposition to associate a CS with a US that follows predictably and immediately is adaptive. Causes often immediately precede effects, so we may associate an effect with the event that preceded it. As we saw in the taste-
Nausea conditioning in cancer patients
Limits on Operant Conditioning
As with classical conditioning, nature sets limits on each species’ capacity for operant conditioning. Science fiction writer Robert Heinlein (1907–
“Once bitten, twice shy.”
G. F. Northall, Folk-Phrases, 1894
We most easily learn and retain behaviors that reflect our biological predispositions. Thus, using food as a reinforcer, you could easily condition a hamster to dig or to rear up, because these are among the animal’s natural food-

In their early days of training animals, psychologists Marian and Keller Breland presumed that operant principles would work on almost any response an animal could make. But along the way, they too learned about biological constraints. In one act, pigs trained to pick up large wooden “dollars” and deposit them in a piggy bank began to drift back to their natural ways. They dropped the coin, pushed it with their snouts as pigs are prone to do, picked it up again, and then repeated the sequence—
304