13.4 Eukaryotic Transcription Is Similar to Bacterial Transcription but Has Some Important Differences
Transcription in eukaryotes is similar to bacterial transcription in that it includes initiation, elongation, and termination, and the basic principles of transcription already outlined apply to eukaryotic transcription. However, there are some important differences. Eukaryotic cells possess three different RNA polymerases, each of which transcribes a different class of RNA and recognizes a different type of promoter. Thus, a generic promoter cannot be described for eukaryotic cells; rather, a promoter’s description depends on whether the promoter is recognized by RNA polymerase I, II, or III. Another difference is in the nature of promoter recognition and initiation. Many proteins take part in the binding of eukaryotic RNA polymerases to DNA templates, and the different types of promoters require different proteins.
Transcription and Nucleosome Structure
Transcription requires sequences on DNA to be accessible to RNA polymerase and other proteins. However, in eukaryotic cells, DNA is complexed with histone proteins in highly compressed chromatin (see Figure 11.4). How can the proteins necessary for transcription gain access to eukaryotic DNA when it is complexed with histones?
The answer to this question is that chromatin structure is modified before transcription so that the DNA is in a more open configuration and is more accessible to the transcription machinery. Several types of proteins have roles in chromatin modification. Acetyltransferases add acetyl groups to amino acids at the ends of the histone proteins, which destabilizes nucleosome structure and makes the DNA more accessible. Other types of histone modification also can affect chromatin packing. In addition, proteins called chromatin-remodeling proteins may bind to the chromatin and displace nucleosomes from promoters and other regions important for transcription. We will take a closer look at the role of changes in chromatin structure associated with gene expression in Chapter 17.
CONCEPTS
The initiation of transcription requires modification of chromatin structure so that DNA is accessible to the transcriptional machinery.
Promoters
A significant difference between bacterial and eukaryotic transcription is the existence of three different eukaryotic RNA polymerases, which recognize different types of promoters. In bacterial cells, the holoenzyme (RNA polymerase plus the sigma factor) recognizes and binds directly to sequences in the promoter. In eukaryotic cells, promoter recognition is carried out by accessory proteins that bind to the promoter and then recruit a specific RNA polymerase (I, II, or III) to the promoter.
One class of accessory proteins comprises general transcription factors, which, along with RNA polymerase, form the basal transcription apparatus—a group of proteins that assemble near the start site and are sufficient to initiate minimal levels of transcription. Another class of accessory proteins consists of transcriptional activator proteins, which bind to specific DNA sequences and bring about higher levels of transcription by stimulating the assembly of the basal transcription apparatus at the start site.
We will focus our attention on promoters recognized by RNA polymerase II, which transcribes the genes that encode proteins. A promoter for a gene transcribed by RNA polymerase II typically consists of two primary parts: the core promoter and the regulatory promoter.
Core Promoter
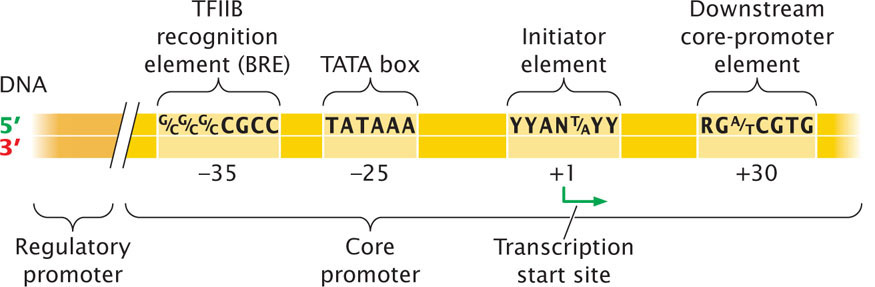
The core promoter is located immediately upstream of the gene (Figure 13.15) and is the site to which the basal transcription apparatus binds. The core promoter typically includes one or more consensus sequences. One of the most common of these sequences is the TATA box, which has the consensus sequence TATAAA and is located from −25 to −30 bp upstream of the start site. Additional consensus sequences that may be found in the core promoters of genes transcribed by RNA polymerase II are shown in Figure 13.15. These consensus sequences are recognized by transcription factors that bind to them and serve as a platform for the assembly of the basal transcription apparatus.
371
Regulatory Promoter
The regulatory promoter is located immediately upstream of the core promoter. A variety of different consensus sequences can be found in the regulatory promoters, and they can be mixed and matched in different combinations. Transcriptional activator proteins bind to these sequences and either directly or indirectly make contact with the basal transcription apparatus and affect the rate at which transcription is initiated. Transcriptional activator proteins also regulate transcription by binding to more-distant sequences called enhancers. The DNA between an enhancer and the promoter loops out, and so transcriptional activator proteins bound to the enhancer can interact with the basal transcription machinery at the core promoter. Enhancers will be discussed in more detail in Chapter 17.
Polymerase I and III Promoters
RNA polymerase I and RNA polymerase III each recognize promoters that are distinct from those recognized by RNA polymerase II. For example, promoters for small rRNA and tRNA genes, transcribed by RNA polymerase III, contain internal promoters that are downstream of the start site and are transcribed into the RNA.
CONCEPTS
General transcription factors and RNA polymerase assemble into the basal transcription apparatus, which binds to DNA near the start site and is necessary for transcription to take place at minimal levels. Additional proteins called transcriptional activators bind to other consensus sequences in promoters and enhancers and affect the rate of transcription.
 CONCEPT CHECK 7
CONCEPT CHECK 7
What is the difference between the core promoter and the regulatory promoter?
- Only the core promoter has consensus sequences.
- The regulatory promoter is farther upstream of the gene.
- Transcription factors bind to the core promoter; transcriptional activator proteins bind to the regulatory promoter.
- Both b and c.
Initiation
Transcription in eukaryotes is initiated through the assembly of the transcriptional machinery on the promoter. This machinery consists of RNA polymerase II and a series of transcription factors that form a giant complex consisting of 50 or more polypeptides. Assembly of the transcription machinery begins when regulatory proteins bind DNA near the promoter and modify the chromatin structure so that transcription can take place. These proteins and other regulatory proteins then recruit the basal transcriptional apparatus to the core promoter.
The basal transcription apparatus consists of RNA polymerase, a series of general transcription factors, and a complex of proteins known as the mediator (Figure 13.16). The general transcription factors include TFIIA, TFIIB, TFIID, TFIIE, TFIIF, and TFIIH, in which TFII stands for transcription factor for RNA polymerase II and the final letter designates the individual factor.
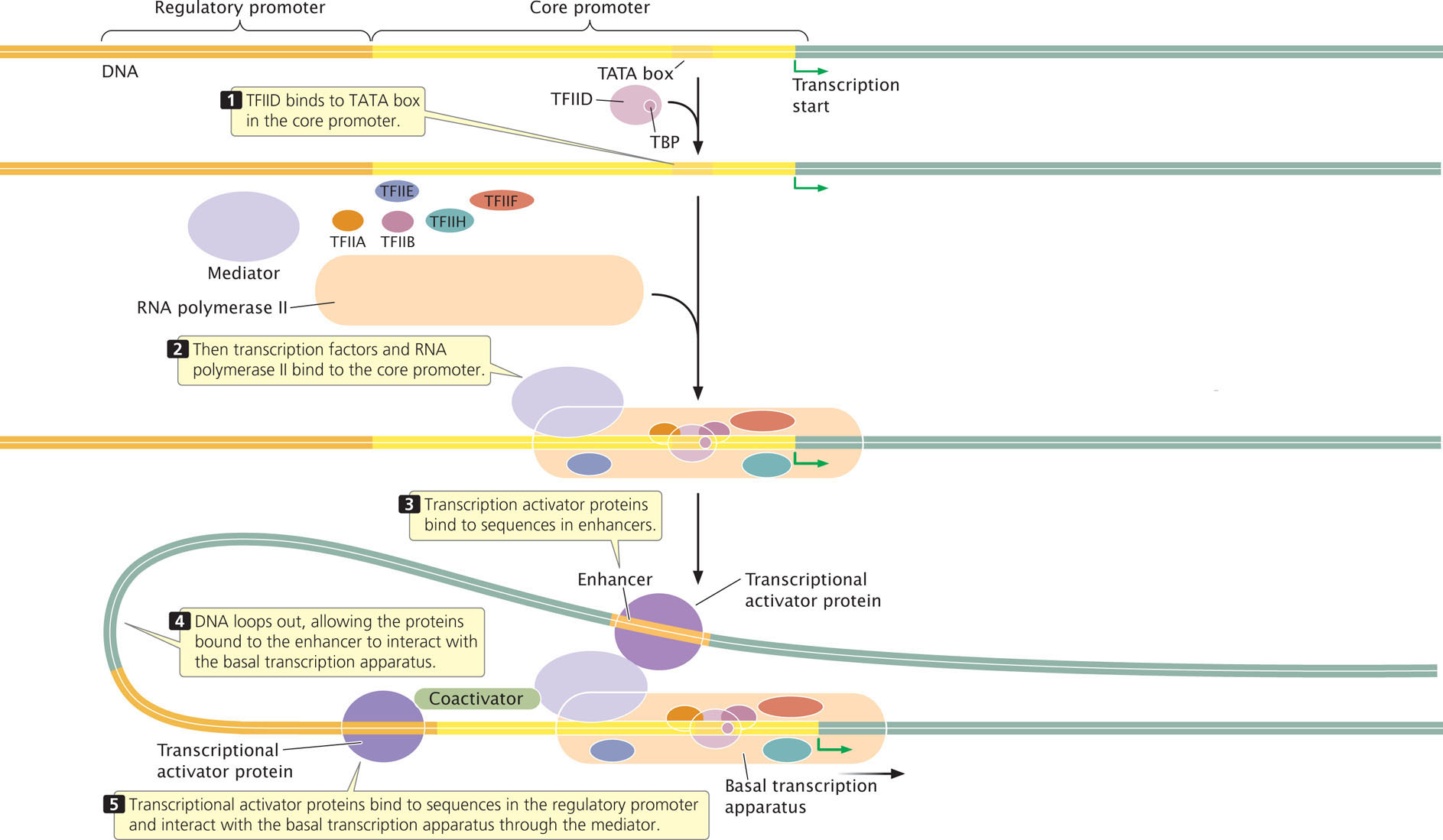
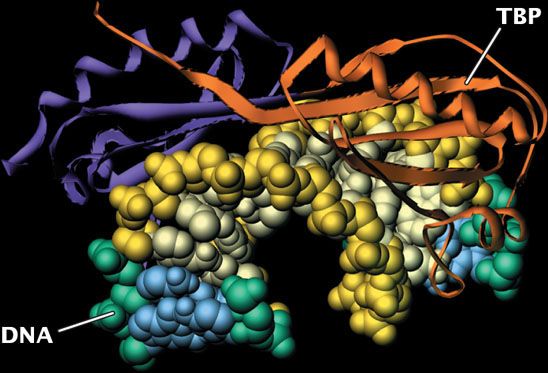
RNA polymerase II and the general transcription factors assemble at the core promoter, forming a pre-initiation complex that is analogous to the closed complex seen in bacterial initiation. Recall that, in bacteria, the sigma factor recognizes and binds to the promoter sequence. In eukaryotes, the function of sigma is replaced by that of the general transcription factors. A first step in initiation is the binding of TFIID to the TATA box on the DNA template. TFIID consists of at least nine polypeptides. One of them is the TATA-binding protein (TBP), which recognizes and binds to the TATA consensus sequence. The TATA-binding protein binds to the minor groove and straddles the DNA as a molecular saddle (Figure 13.17), bending the DNA and partly unwinding it. Other transcription factors bind to additional consensus sequences in the core promoter and to RNA polymerase and position the polymerase over the transcription start site.
372
After the RNA polymerase and transcription factors have assembled on the core promoter, conformational changes take place in both the DNA and the polymerase. These changes cause from 11 to 15 bp of DNA surrounding the transcription start site to separate, producing the single-stranded DNA that will serve as a template for transcription.
The single-stranded DNA template is positioned within the active site of RNA polymerase, creating a structure called the open complex. After the open complex has formed, the synthesis of RNA begins as phosphate groups are cleaved off nucleoside triphosphates and nucleotides are joined together to form an RNA molecule. As in bacterial transcription, RNA polymerase may generate and release several short RNA molecules in abortive transcription before the polymerase initiates the synthesis of a full-length RNA molecule. ![]() TRY PROBLEM 35
TRY PROBLEM 35
373
CONCEPTS
Transcription is initiated when the basal transcription apparatus, consisting of RNA polymerase and transcription factors, assembles on the core promoter and becomes an open complex.
CONCEPT CHECK 8
What is the role of TFIID in transcription initiation?
Elongation
After about 30 bp of RNA have been synthesized, the RNA polymerase leaves the promoter and enters the elongation stage of transcription. Many of the transcription factors are left behind at the promoter and can serve to quickly reinitiate transcription with another RNA polymerase enzyme.
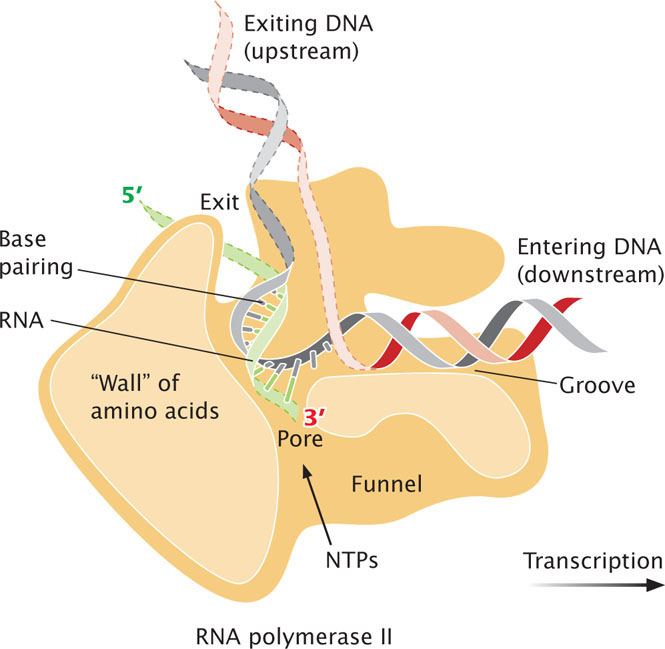
The molecular structure of eukaryotic RNA polymerase II and how it functions during elongation have been revealed through the work of Roger Kornberg and his colleagues, for which Kornberg was awarded a Nobel Prize in chemistry in 2006. The RNA polymerase maintains a transcription bubble during elongation, in which about eight nucleotides of RNA remain base paired with the DNA template strand. The DNA double helix enters a cleft in the polymerase and is gripped by jawlike extensions of the enzyme (Figure 13.18). The two strands of the DNA are unwound and RNA nucleotides that are complementary to the template strand are added to the growing 3' end of the RNA molecule. As it funnels through the polymerase, the DNA-RNA hybrid hits a wall of amino acids and bends at almost a right angle; this bend positions the end of the DNA-RNA hybrid at the active site of the polymerase, and new nucleotides are added to the 3' end of the growing RNA molecule. The newly synthesized RNA is separated from the DNA and runs through another groove before exiting from the polymerase.
Termination
The three eukaryotic RNA polymerases use different mechanisms for termination. RNA polymerase I requires a termination factor like the rho factor utilized in the termination of some bacterial genes. Unlike rho, which binds to the newly transcribed RNA molecule, the termination factor for RNA polymerase I binds to a DNA sequence downstream of the termination site.
RNA polymerase III ends transcription after transcribing a terminator sequence that produces a string of uracil nucleotides in the RNA molecule, like that produced by the rho-independent terminators of bacteria. Unlike rho-independent terminators in bacterial cells, however, RNA polymerase III does not require that a hairpin structure precede the string of Us.
The termination of transcription by RNA polymerase II is not at specific sequences. Instead, RNA polymerase II often continues to synthesize RNA hundreds or even thousands of nucleotides past the coding sequence necessary to produce the mRNA. As we will see in Chapter 14, the end of pre-mRNA is cleaved at a specific site, designated by a consensus sequence, while transcription is still taking place at the 3′ end of the molecule. Cleavage cuts the pre-mRNA into two pieces: the mRNA that will eventually encode the protein and another piece of RNA that has its 5′ end trailing out of the RNA polymerase (Figure 13.19). An enzyme (called Rat1 in yeast) attaches to the 5′ end of this RNA and moves toward the 3 end where RNA polymerase continues the transcription of RNA. Rat1 is a 5′→3′ exonuclease−an enzyme capable of degrading RNA in the 5′→3′ direction. Like a guided torpedo, Rat1 homes in on the polymerase, chewing up the RNA as it moves. When Rat1 reaches the transcriptional machinery, transcription terminates. Note that this mechanism is similar to that of rho-dependent termination in bacteria (see Figure 13.13), except that rho does not degrade the RNA molecule.
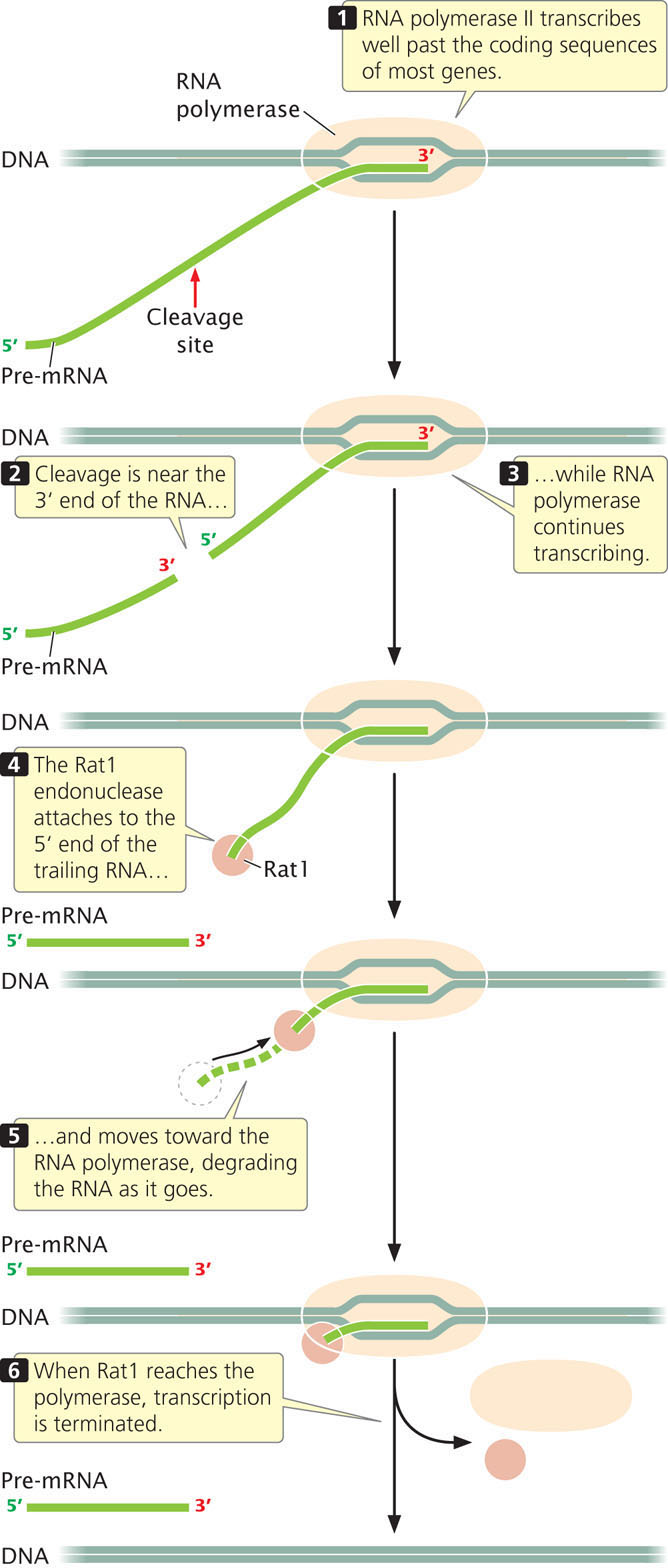
CONCEPTS
The different eukaryotic RNA polymerases use different mechanisms of termination. Transcription at genes transcribed by RNA polymerase II is terminated when an exonuclease enzyme attaches to the cleaved 5′ end of the RNA, moves down the RNA, and reaches the polymerase enzyme.
CONCEPT CHECK 9
How are the processes of RNA polymerase II termination and rho-dependent termination in bacteria similar and how are they different?
374