The Origin of Spineless Sticklebacks
633

Threespine sticklebacks (Gasterosteus aculeatus) are curious little fish. In spite of their small size—they reach only about 2 inches (5 cm) in length—they are heavily armored, with protective plates on their back, sides, and belly, and possess three spines on their dorsal surface, thus giving rise to their name. Each fish also possesses two impressive pelvic spines, which are anchored to the pelvic girdle and project out from their sides (Figure 22.1). The dorsal and pelvic spines make sticklebacks difficult to swallow, allowing them to survive in an environment where an unprotected 2-inch fish is an easily caught meal for numerous predators.
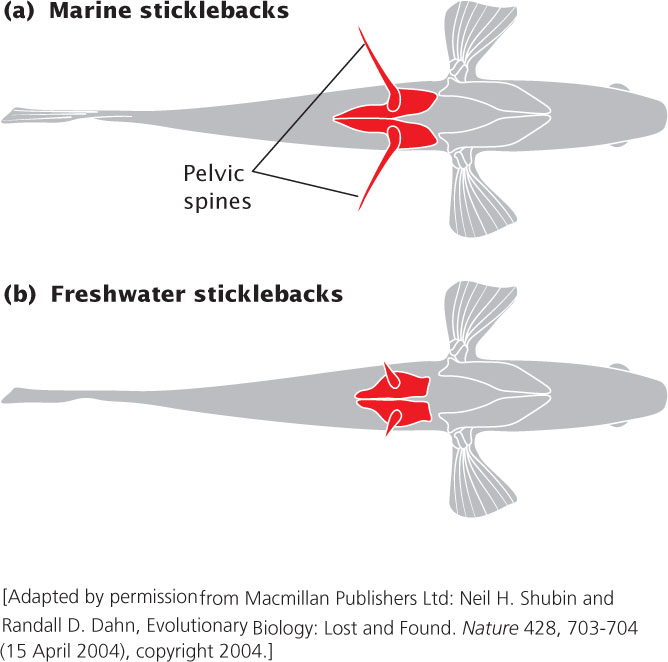
Most sticklebacks are marine, living in the ocean, but a few isolated populations can be found inland in freshwater lakes. In North America, these freshwater populations originated 10,000 to 20,000 years ago, at the end of the last ice age, when marine sticklebacks invaded the lakes. Many lake populations of threespine sticklebacks have lost their armor and spines, probably because fish predators that might eat them are absent, there is little calcium present to develop the plates and spines, and invertebrate predators found in the lakes catch the fish by grabbing onto their spines. Biologists have long been interested in how marine sticklebacks made the evolutionary transition from the marine to the freshwater environment: how did a heavily armored fish become spineless? Research conducted by developmental geneticists has begun to provide an answer to this question.
In 1998, geneticist David Kingsley from Stanford University began a collaboration with Dolph Schluter, an evolutionary biologist from the University of British Columbia. Their goal was to understand how threespine sticklebacks lost their pelvic spines during the evolutionary transition from marine to freshwater environments. The scientists crossed a female marine stickleback that possessed spines with a male from Paxton Lake, British Columbia that lacked pelvic spines. All of the F1 fish from this cross possessed pelvic spines. They then crossed two of the F1 fish, producing a total of 375 F2 progeny. These F2 progeny showed a wide range of variation in their pelvic spines: some had fully developed spines, some lacked spines altogether, and others had varying degrees of spine reduction.
Kingsley and his colleagues then examined the association of pelvic spines in the F2 progeny and the inheritance of genetic markers across the genome. They found that most of the variation in pelvic spines was associated with genetic markers from a particular region on chromosome 7. Interestingly, this same region contains Pitx1, a gene that is often expressed in the hind limbs of developing vertebrates. Mice with a mutation in their Pitx1 gene often have reduced hind limbs, as well as other developmental abnormalities. This observation suggested that mutations within the Pitx1 gene might be responsible for the absence of pelvic spines in freshwater populations of sticklebacks. But when the researchers examined the DNA sequences of the Pitx1 gene in marine and freshwater threespine sticklebacks, they found no differences that would alter the amino acid sequence of the protein encoded by the gene. What they did find was that the expression of the Pitx1 gene differed in marine and freshwater fish. Sticklebacks with pelvic fins expressed Pitx1 in their pelvic region; sticklebacks without pelvic fins expressed Pitx1 in other tissues, but the gene was completely inactive in the pelvic region. This suggested that although the Pitx1 gene itself is not mutated in spineless fish, mutations in regulatory elements that affect the expression of Pitx1 might be the source of variation in presence or absence of pelvic spines.
634
In 2010, Kingsley and his team located the mutation that causes the absence of spines in freshwater populations of threespine sticklebacks. They found an enhancer 500 bp upstream of the Pitx1 gene that controls the tissue-specific expression of Pitx1. Enhancers are DNA sequences that promote the transcription of distant genes, often in a tissue-specific manner (see Chapter 17). The researchers determined that spineless fish from Paxton Lake possessed a deletion that removed this enhancer, thus preventing expression of Pitx1 and ultimately resulting in the loss of pelvic spines. Spineless fish from other lakes in Canada, Alaska, and even Iceland also possessed deletions of the enhancer, but fish from different lakes possessed different deletions. This observation suggests that during the course of evolution, pelvic spines have been lost multiple times through natural selection acting on different mutations that have the same phenotypic effect.
The story of how the stickleback lost its spines illustrates that major anatomical alterations can occur through small genetic changes in key regulatory sequences that affect development, a theme throughout this chapter on the genetic control of development. The chapter begins with a consideration of how cell differentiation occurs, not through loss of genes but rather through alteration of gene expression. We then discuss the genetic control of early development of Drosophila embryos, one of the best-understood developmental systems. We next consider the genetic control of floral structure in plants, another model system that has been well studied, followed by a more detailed look at programmed cell death and the use of development for understanding evolution. At the end of the chapter, we turn to the development of immunity and its genetic control.