26.3 New Species Arise Through the Evolution of Reproductive Isolation
Earlier we discussed two types of evolution: anagenesis (change within a lineage), and cladogenesis, the splitting of lineages. Cladogenesis arises through speciation, the process by which one population separates into two distinct evolutionary groups.
The term species literally means kind or appearance; species are different kinds or types of living organisms. In many cases species differences are easy to recognize: a horse is clearly a different species from a chicken. Sometimes, however, species differences are not so clear. Some species of Plethodon salamanders are so similar in appearance that they can be distinguished only by looking at their proteins or genes.
The concept of a species has two primary uses in biology. First, species is a name given to a particular type of organism. For effective communication, biologists must use a standard set of names for the organisms that they study, and species names serve that purpose. When a geneticist talks about conducting crosses with Drosophila melanogaster, other biologists immediately understand which organism was used. The second use of the term species is in an evolutionary context: a species is considered an evolutionarily independent group of organisms.
The Biological Species Concept
What kinds of differences are required to consider two organisms different species? A widely used definition of species is the biological species concept, first fully developed by evolutionary biologist Ernst Mayr in 1942. Mayr was primarily interested in the biological characteristics that are responsible for separating organisms into independently evolving units. He defined a species as a group of organisms whose members are capable of interbreeding with one another but are reproductively isolated from the members of other species. In other words, members of the same species have the biological potential to exchange genes, and members of different species cannot exchange genes. Because different species do not exchange genes, each species evolves independently.
750
Not all biologists adhere to the biological species concept, and there are several problems associated with it. For example, reproductive isolation, on which the biological species concept is based, cannot be determined from fossils and, in practice it is often difficult to determine whether even living species are biologically capable of exchanging genes. Furthermore, the biological species concept cannot be applied to asexually reproducing organisms, such as bacteria. In practice, most species are distinguished on the basis of phenotypic (usually anatomical) differences. Biologists often assume that phenotypic differences represent underlying genetic differences; if the phenotypes of two organisms are quite different, then they probably cannot and do not interbreed in nature. Because of these problems, some biologists have proposed alternative definitions for a species. For example, the morphospecies concept defines a species based entirely on phenotypic (morphological) similarities and differences. The phylogenetic species concept defines a species as the smallest recognizable group that has a unique evolutionary history. Here we will use the biological species concept because it is widely used and is based on genetic differences.
Reproductive Isolating Mechanisms
The key to species differences under the biological species concept is reproductive isolation—biological characteristics that prevent genes from being exchanged between different species. Any biological factor or mechanism that prevents gene exchange is termed a reproductive isolating mechanism.
Prezygotic Reproductive Isolating Mechanisms
Some species are separated by prezygotic reproductive isolating mechanisms, which prevent gametes from two different species from fusing and forming a hybrid zygote. In ecological isolation, members of two species do not encounter one another and therefore do not reproduce with one another: they have different ecological niches, living in different habitats and interacting with the environment in different ways. For example, some species of forest-dwelling birds feed and nest in the forest canopy, whereas others confine their activities to the forest floor. Because they never come into contact, these birds are reproductively isolated from one another. Other species are separated by behavioral isolation, differences in behavior that prevent interbreeding. Many male frogs attract females of the same species by using a unique, species-specific call. Two closely related frogs may use the same pond but never interbreed, because females are attracted only to the call of their own species.
Another type of prezygotic reproductive isolation is temporal isolation, in which reproduction takes place at different times of the year. Some species of plants do not exchange genes, because they flower at different times of the year. Mechanical isolation results from anatomical differences that prevent successful copulation. This type of isolation is seen in many insects, in which closely related species differ in their male and female genitalia, and so copulation is physically impossible. Finally, some species are separated by gametic isolation, in which mating between individuals of different species takes place, but the gametes do not form zygotes. Male gametes may not survive in the female reproductive tract or may not be attracted to female gametes. In other cases, male and female gametes meet but are too incompatible to fuse to form a zygote. Gametic isolation is seen in many plants, where pollen from one species cannot fertilize the ovules of another species.
Postzygotic Reproductive Isolating Mechanisms
Other species are separated by postzygotic reproductive isolating mechanisms, in which gametes of two species fuse and form a zygote, but there is no gene flow between the two species, either because the resulting hybrids are inviable or sterile or because reproduction breaks down in subsequent generations.
If prezygotic reproductive isolating mechanisms fail or have not yet evolved, mating between two organisms of different species may take place, with the formation of a hybrid zygote containing genes from two different species. In many cases, such species are still separated by hybrid inviability, in which incompatibility between genomes of the two species prevents the hybrid zygote from developing. Hybrid inviability is seen in some groups of frogs, in which mating between different species and fertilization take place, but the resulting embryos never complete development.
Other species are separated by hybrid sterility, in which hybrid embryos complete development but are sterile, so that genes are not passed between species. Donkeys and horses frequently mate and produce a viable offspring—a mule—but most mules are sterile; thus, there is no gene flow between donkeys and horses (but see Problem 47 at the end of Chapter 8). Finally, some closely related species are capable of mating and producing viable and fertile F1 progeny. However, genes do not flow between the two species because of hybrid breakdown, in which further crossing of the hybrids produces inviable or sterile offspring. The different types of reproductive isolating mechanisms are summarized in Table 26.2.
| Type | Characteristics |
|---|---|
| Prezygotic | Before a zygote has formed |
| Ecological | Differences in habitat; individuals do not meet |
| Temporal | Reproduction takes place at different times |
| Mechanical | Anatomical differences prevent copulation |
| Behavioral | Differences in mating behavior prevent mating |
| Gametic | Gametes incompatible or not attracted to each other |
| Postzygotic | After a zygote has formed |
| Hybrid inviability | Hybrid zygote does not survive to reproduction |
| Hybrid sterility | Hybrid is sterile |
| Hybrid breakdown | F1 hybrids are viable and fertile, but F2 are inviable or sterile |
CONCEPTS
The biological species concept defines a species as a group of potentially interbreeding organisms that are reproductively isolated from the members of other species. Under this concept, species are separated by prezygotic or postzygotic reproductive isolating mechanisms.
 CONCEPT CHECK 4
CONCEPT CHECK 4
Which statement is an example of postzygotic reproductive isolation?
- Sperm of species A dies in the oviduct of species B before fertilization can take place.
- Hybrid zygotes between species A and B are spontaneously aborted early in development.
- The mating seasons of species A and B do not overlap.
- Males of species A are not attracted to the pheromones produced by the females of species B.
Modes of Speciation
Speciation is the process by which new species arise. In regard to the biological species concept, speciation comes about through the evolution of reproductive isolating mechanisms.
There are two principle ways in which new species arise. Allopatric speciation arises when a geographic barrier first splits a population into two groups and blocks the exchange of genes between them. The interruption of gene flow then leads to the evolution of genetic differences that result in reproductive isolation. Sympatric speciation is speciation that arises in the absence of any external barrier to gene flow; reproductive isolating mechanisms evolve within a single population. We will take a more detailed look at both of these mechanisms next.
Allopatric Speciation
Allopatric speciation is initiated when a geographic barrier splits a population into two or more groups and prevents gene flow between the isolated groups (Figure 26.6a). Geographic barriers can take a number of forms. Uplifting of a mountain range may split a population of lowland plants into separate groups on each side of the mountains. Oceans serve as effective barriers for many types of terrestrial organisms, separating individuals on different islands from one another and from those on the mainland. Rivers often separate populations of fish located in separate drainages. The erosion of mountains may leave populations of alpine plants isolated on separate mountain peaks.
After two populations have been separated by a geographic barrier that prevents gene flow between them, they evolve independently (Figure 26.6b). The genetic isolation allows each population to accumulate genetic differences that are not found in the other population; genetic differences arise through natural selection, unique mutations, and genetic drift (if the populations are small). Genetic differentiation may eventually lead to prezygotic and postzygotic isolation. It is important to note that prezygotic isolation and postzygotic isolation arise simply as a consequence of genetic divergence.
If the geographic barrier that once separated the two populations disappears or individuals are able to disperse over it, the populations come into secondary contact (Figure 26.6c). At this point, several outcomes are possible. If limited genetic differentiation has taken place during the separation of the populations, reproductive isolating mechanisms may not have evolved or may be incomplete. Genes will flow between the two populations, eliminating any genetic differences that did arise, and the populations will remain a single species.
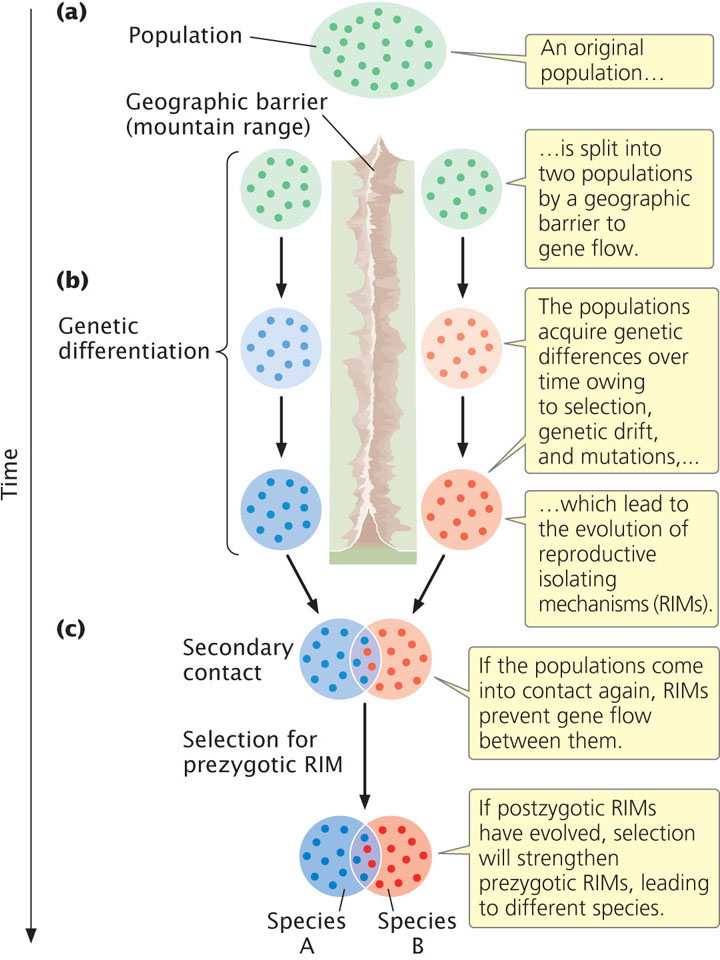
A second possible outcome is that genetic differentiation during separation leads to prezygotic reproductive isolating mechanisms; in this case, the two populations are different species. A third possible outcome is that, during their time apart, some genetic differentiation took place between the populations, leading to incompatibility in their genomes and postzygotic isolation. If postzygotic isolating mechanisms have evolved, any mating between individuals from the different populations will produce hybrid offspring that are inviable or sterile. Individuals that mate only with members of the same population will have higher fitness than that of individuals that mate with members of the other population; so natural selection will increase the frequency of any trait that prevents interbreeding between members of the different populations. With the passage of time, prezygotic reproductive isolating mechanisms will evolve. In short, if some postzygotic reproductive isolation exists, natural selection will favor the evolution of prezygotic reproductive isolating mechanisms to prevent wasted reproduction by individuals mating with members of the other population. This process of postzygotic reproductive isolation leading to the evolution of prezygotic isolating mechanisms is termed reinforcement.
752
A number of variations in this general model of allopatric speciation are possible. Many new species probably arise when a small group of individuals becomes geographically isolated from the main population; for example, a few individuals of a mainland population might migrate to a geographically isolated island. In this situation, founder effect and genetic drift play a larger role in the evolution of genetic differences between the populations.
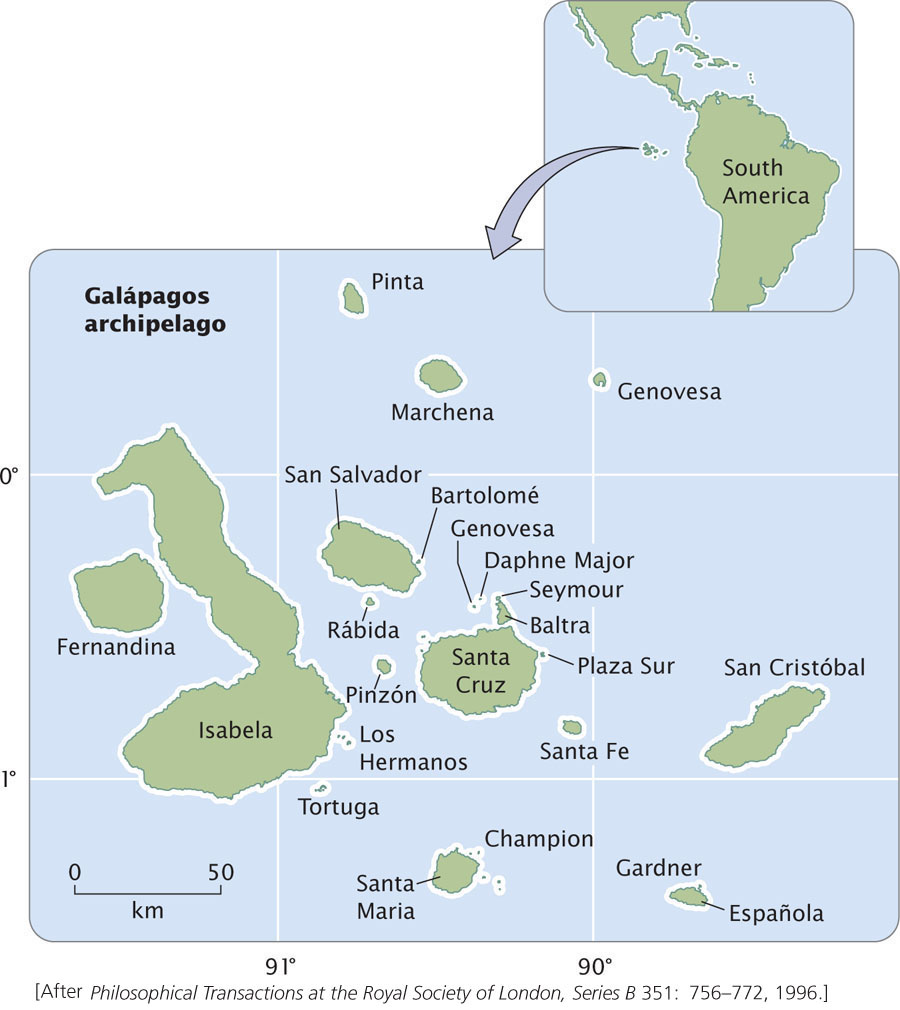
An excellent example of allopatric speciation can be found in Darwin’s finches, a group of birds that Charles Darwin discovered on the Galápagos Islands during his voyage aboard the Beagle. The Galápagos are an archipelago of islands located about 900 km off the coast of South America (Figure 26.7). Consisting of more than a dozen large islands and many smaller ones, the Galápagos formed from volcanoes that erupted over a geological hot spot that has remained stationary while the geological plate over it moved eastward in the past 3 million years. The movement of the geological plate pulled newly formed islands eastward, and so the islands to the east (San Cristóbal and Española) are older than those to the west (Isabela and Fernandina). With the passage of time, the number of islands in the archipelago increased as new volcanoes arose.
Darwin’s finches consist of 14 species that are found on various islands in the Galápagos archipelago (Figure 26.8). The birds vary in the shape and sizes of their beaks, which are adapted for eating different types of food items. Recent studies of the development of finch embryos have helped to reveal some of the molecular details of how differences in beak shapes have evolved.
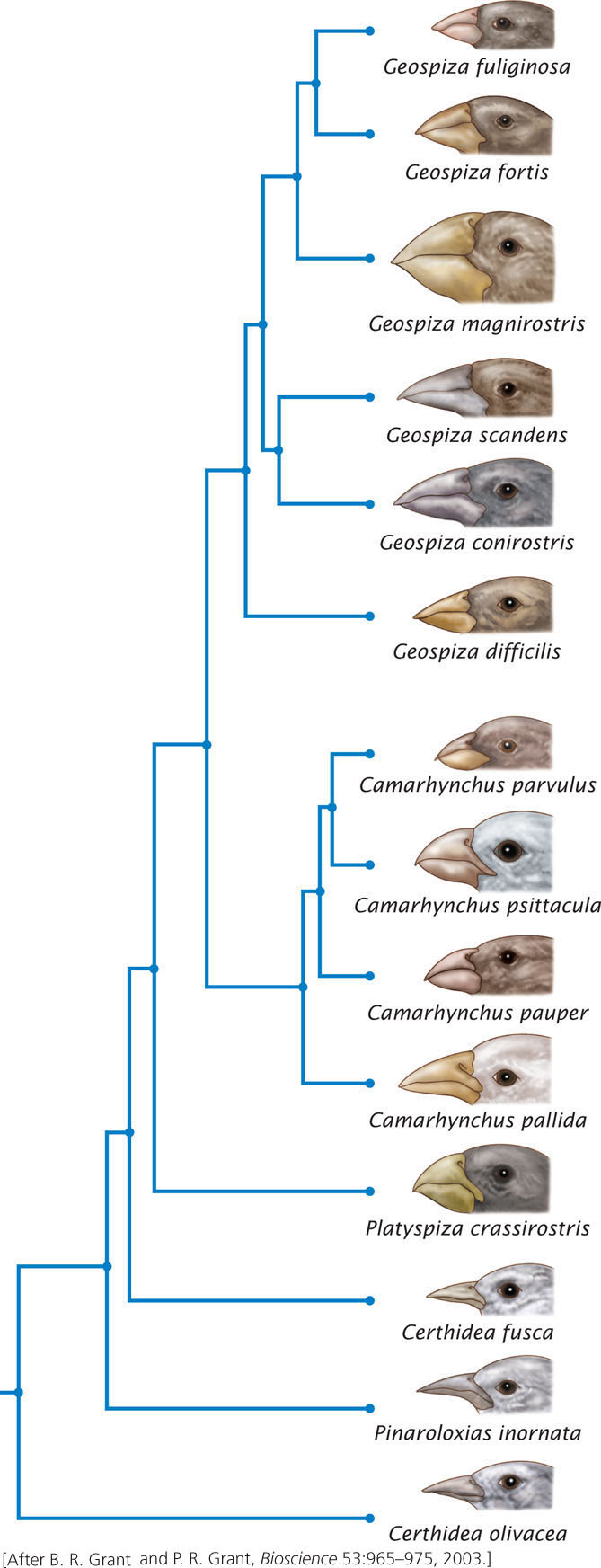
Genetic studies have demonstrated that all the birds are closely related and evolved from a single ancestral species that migrated to the islands from the coast of South America some 2 million to 3 million years ago. The evolutionary relationships among the 14 species, based on studies of microsatellite data, are depicted in Figure 26.8. Most of the species are separated by a behavioral isolating mechanism (song in particular), but some of the species can and occasionally do hybridize in nature.
The first finches to arrive in the Galápagos probably colonized one of the larger eastern islands. A breeding population became established and increased with time. At some point, a few birds dispersed to other islands, where they were effectively isolated from the original population, and established a new population. The new population underwent genetic differentiation owing to genetic drift and adaptation to the local conditions of the island, and it eventually became reproductively isolated from the original population. Individual birds from the new population then dispersed to other islands and gave rise to additional species. This process was repeated many times. Occasionally, newly evolved birds dispersed to an island where another species was already present, giving rise to secondary contact between the species. Today, many of the islands have more than one resident finch.
753
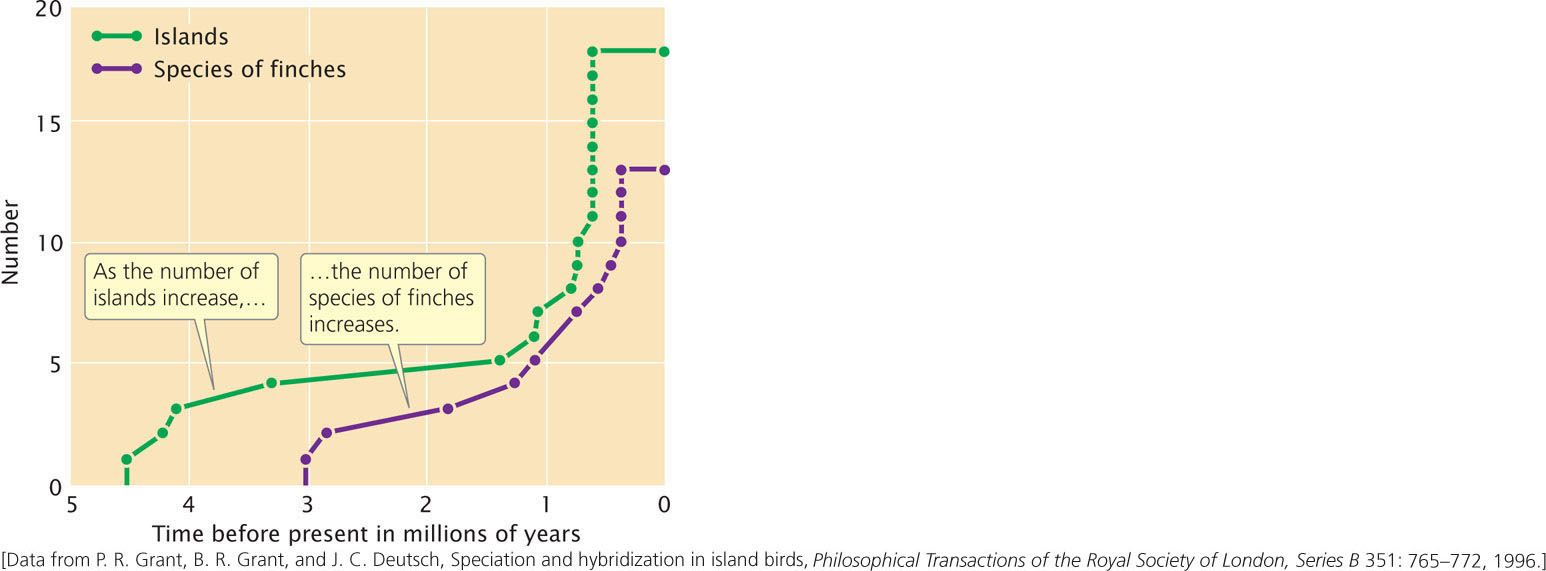
The age of the 14 species has been estimated with data from mitochondrial DNA. Figure 26.9 shows that there is a strong correspondence between the number of bird species present at various times in the past and the number of islands in the archipelago. This correspondence is one of the most compelling pieces of evidence for the theory that the different species of finches arose through allopatric speciation.
CONCEPTS
Allopatric speciation is initiated when a geographic barrier to gene flow splits a single population into two or more populations. With the passage of time, the populations evolve genetic differences, which bring about reproductive isolation. After postzygotic reproductive isolating mechanisms have evolved, selection favors the evolution of prezygotic reproductive isolating mechanisms.
CONCEPT CHECK 5
What role does genetic drift play in allopatric speciation?
Sympatric Speciation
Sympatric speciation arises in the absence of any geographic barrier to gene flow; reproductive isolating mechanisms evolve within a single interbreeding population. Sympatric speciation has long been controversial within evolutionary biology. Ernst Mayr believed that sympatric speciation was impossible, and he demonstrated that many apparent cases of sympatric speciation could be explained by allopatric speciation. More recently, however, evidence has accumulated that sympatric speciation can and has arisen under special circumstances. The difficulty with sympatric speciation is that isolating mechanisms arise as a consequence of genetic differentiation, which takes place only if gene flow between groups is interrupted. But, without reproductive isolation (or some external barrier), how can gene flow be interrupted? How can genetic differentiation arise within a single group that is freely exchanging genes?
754
Most models of sympatric speciation assume that genetic differentiation is initiated by strong disruptive selection taking place within a single population. One homozygote (A1A1) is strongly favored on one resource (perhaps the plant species that is host to an insect) and the other homozygote (A2A2) is favored on a different resource (perhaps a different host plant). Heterozygotes (AA2) have low fitness on both resources. In this situation, natural selection will favor genotypes at other loci that cause assortative mating (matings between like individuals, see Chapter 25), and so no matings take place between A1A1 and A2A2, which would produce A1A2 offspring with low fitness.
Now imagine that alleles at a second locus affect mating behavior, such that C1C1 individuals prefer mating only with other C1C1 individuals, and C2C2 individuals prefer mating with other C2C2 individuals. If alleles at the A locus are non-randomly associated with alleles at the C locus so that only A1A1 C1C1 individuals and A2A2 C2C2 individuals exist, then gene flow will be restricted between individuals using the different resources, allowing the two groups to evolve further genetic differences that might lead to reproductive isolation and sympatric speciation.
The difficulty with this model is that recombination quickly breaks up the nonrandom associations between genotypes at the two loci, producing individuals such as A1A1 C2C2, which would prefer to mate with A2A2 C2C2 individuals. This mating would produce all A1A2 offspring, which do poorly on both resources. Thus, even limited recombination will prevent the evolution of the mating-preference genes.
Sympatric speciation is more probable if the genes that affect resource utilization also affect mating preferences. Genes that affect both resource utilization and mating preference are indeed present in some host races—populations of specialized insects that feed on different host plants. Guy Bush studied what appeared to be initial stages of speciation in host races of the apple maggot fly (Rhagoletis pomonella, Figure 26.10). The flies of this species feed on the fruits of a specific host tree. Mating takes place near the fruits, and the flies lay their eggs on the ripened fruits, where their larvae grow and develop. R. pomonella originally existed only on the fruits of hawthorn trees, which are native to North America; 150 years ago, R. pomonella was first observed on cultivated apples, which are related to hawthorns but a different species. Infestations of apples by this new apple host race of R. pomonella quickly spread, and, today, many apple trees throughout North America are infested with the flies.

The apple host race of R. pomonella probably originated when a few flies acquired a mutation that allowed them to feed on apples instead of hawthorn fruits. Because mating takes place on and near the fruits, flies that use apples are more likely to mate with other flies using apples, leading to genetic isolation between flies using hawthorns and those using apples. Indeed, Bush found that some genetic differentiation has already taken place between the two host races. Flies lay their eggs on ripening fruit, and there has been strong selection for the flies to synchronize their reproduction with the period when their host species has ripening fruit. Apples ripen several weeks earlier than hawthorns. Correspondingly, the peak mating period of the apple host races is 3 weeks earlier than that of the hawthorn race. These differences in the timing of reproduction between apple and hawthorn races have further reduced gene flow—to about 2%—between the two host races and have led to significant genetic differentiation between them. These differences have evolved in the past 150 years and evolution appears to be ongoing. Although genetic differentiation has taken place between apple and hawthorn host races of R. pomonella and some degree of reproductive isolation has evolved between them, reproductive isolation is not yet complete and speciation has not fully taken place. ![]() TRY PROBLEM 22
TRY PROBLEM 22
Speciation Through Polyploidy
A special type of sympatric speciation takes place through polyploidy (see Chapter 8). Polyploid organisms have more than two genomes (3n, 4n, 5n, etc.). As discussed in Chapter 8, allopolyploidy often arises when two diploid species hybridize, producing 2n hybrid offspring. Nondisjunction in one of the hybrid offspring produces a 4n tetraploid. Because this tetraploid contains exactly two copies of each chromosome, it is usually fertile and will be reproductively isolated from the two parental species by differences in chromosome number (see Figure 8.28).
Numerous species of flowering plants are allopolyploids. Speciation through polyploidy was observed when it led to a new species of salt-marsh grass that arose along the coast of England around 1870. This polyploid contains genomes of the European salt grass Spartina maritima (2n = 60) and the American salt grass S. alterniflora (2n = 62; Figure 26.11).
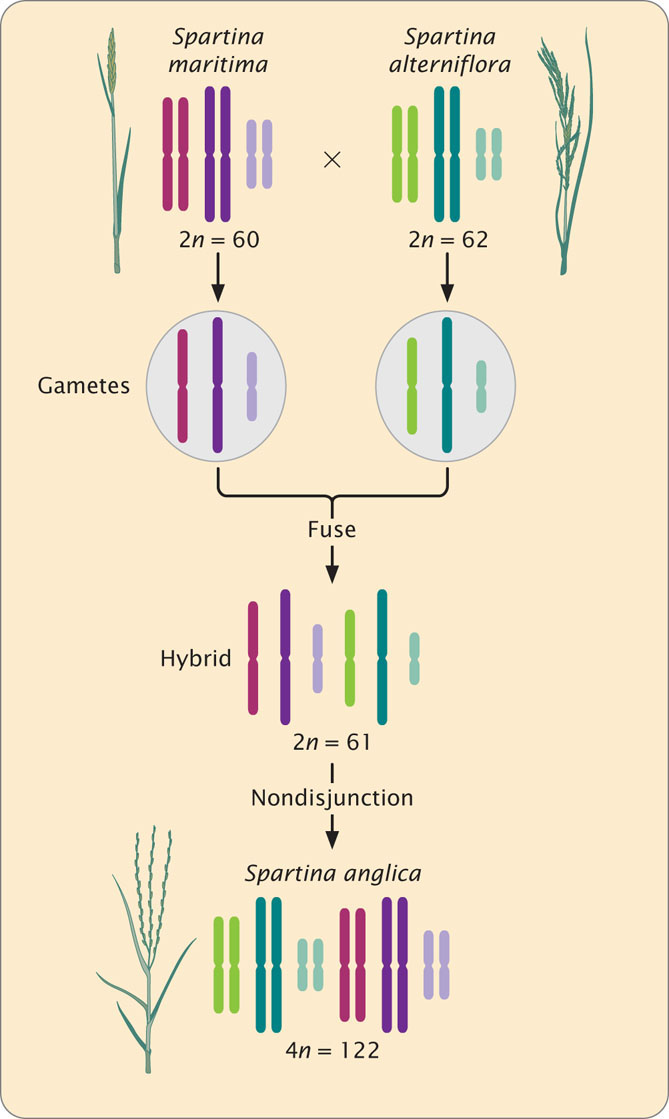
755
Seeds from the American salt grass were probably transported to England in the ballast of a ship. Regardless of how it got there, S. alterniflora grew in an English marsh and eventually crossed with S. maritima, producing a hybrid with 2n = 61. Nondisjunction in the hybrid then led to chromosome doubling, producing a new species S. anglica with 4n = 122 (see Figure 26.11). This new species subsequently spread along the coast of England.
CONCEPTS
Sympatric speciation arises within a single interbreeding population without any geographic barrier to gene flow. Sympatric speciation may arise under special circumstances, such as when resource use is linked to mating preference (in host races) or when species hybridization leads to allopolyploidy.
Genetic Differentiation Associated with Speciation
As we have seen, genetic differentiation leads to the evolution of reproductive isolating mechanisms, which restrict gene flow between populations and lead to speciation. How much genetic differentiation is required for reproductive isolation to take place? This question has received considerable study by evolutionary geneticists, but, unfortunately, there is no universal answer. Some newly formed species differ in many genes, whereas others appear to have undergone divergence in just a few genes.
One group of organisms that has been extensively studied for genetic differences associated with speciation is the genus Drosophila. The Drosophila willistoni group consists of at least 12 species found in Central and South America in various stages in the process of speciation. Using protein electrophoresis, Francisco Ayala and his colleagues genotyped flies from different geographic populations (populations with limited genetic differences), subspecies (populations with considerable genetic differences), sibling species (newly arisen species), and nonsibling species (older species). For each group, they computed a measure of genetic similarity, which ranges from 1 to 0 and represents the overall level of genetic differentiation (Table 26.3). They found that there was a general decrease in genetic similarity as flies evolve from geographic populations to subspecies to sibling species to nonsibling species. These data suggest that considerable genetic differentiation at many loci is required for speciation to arise. A study of D. simulans and D. melanogaster, two species that produce inviable hybrids when crossed, suggested that at least 200 genes contribute to the inviability of hybrids between the two species.
| Group | Mean Genetic Similarity |
|---|---|
| Geographic populations | 0.970 |
| Subspecies | 0.795 |
| Sibling species | 0.517 |
| Nonsibling species | 0.352 |
However, other studies suggest that speciation may have arisen through changes in just a few genes. For example, D. heteroneura and D. silvestris are two species of Hawaiian fruit flies that exhibit behavioral reproductive isolation. The isolation is determined largely by differences in head shape; D. heteroneura has a hammer-shaped head with widely separated eyes that is recognized by females of the same species but rejected by D. silvestris females. Genetic studies indicate that only a few loci (about 10) determine the differences in head shape.
In another study, researchers examined genetic differences in two closely related species of birds, the collared flycatcher (Ficedula albicollis) and pied flycatcher (F hypoleuca), both of which occur in Europe. These two species are believed to have separated from a common ancestor less than 2 million years ago, probably by allopatric speciation associated with glaciation. In areas where both species occur together, collared flycatchers and pied flycatcher will sometimes mate, but the two species are separated by both pre- and postzygotic reproductive isolating mechanisms. For example, females prefer males with the plumage and song of their own species, and hybrids do not survive as well as offspring of parents both from the same species.
756
Researchers at Uppsala University in Sweden completely sequenced the genomes of 10 male birds of each species. They found that large parts of the genomes of the two species were the same: for example, only about 5 out of 1000 nucleotides differed between species, compared with 4 out of 1000 nucleotides within species. The differences that did exist between the species were not uniform across the genome; rather, the differences tended to be concentrated in a few “divergence islands,” which had up to 50 times greater sequence differences than the average for the genome. A few divergence islands were found on each chromosome, and they occurred primarily near centromeres and telomeres, suggesting that differences in chromosome structure play an important role in speciation. The divergence islands tended to show low levels of differences within species, indicating that natural selection had favored different genes in each region.
CONCEPTS
Some newly arising species have a considerable number of genetic differences; others have few genetic differences.