2.2 Cell Reproduction Requires the Copying of the Genetic Material, Separation of the Copies, and Cell Division
For any cell to reproduce successfully, three fundamental events must take place: (1) its genetic information must be copied, (2) the copies of genetic information must be separated from each other, and (3) the cell must divide. All cellular reproduction includes these three events, but the processes that lead to these events differ in prokaryotic and eukaryotic cells because of their structural differences.
Prokaryotic Cell Reproduction
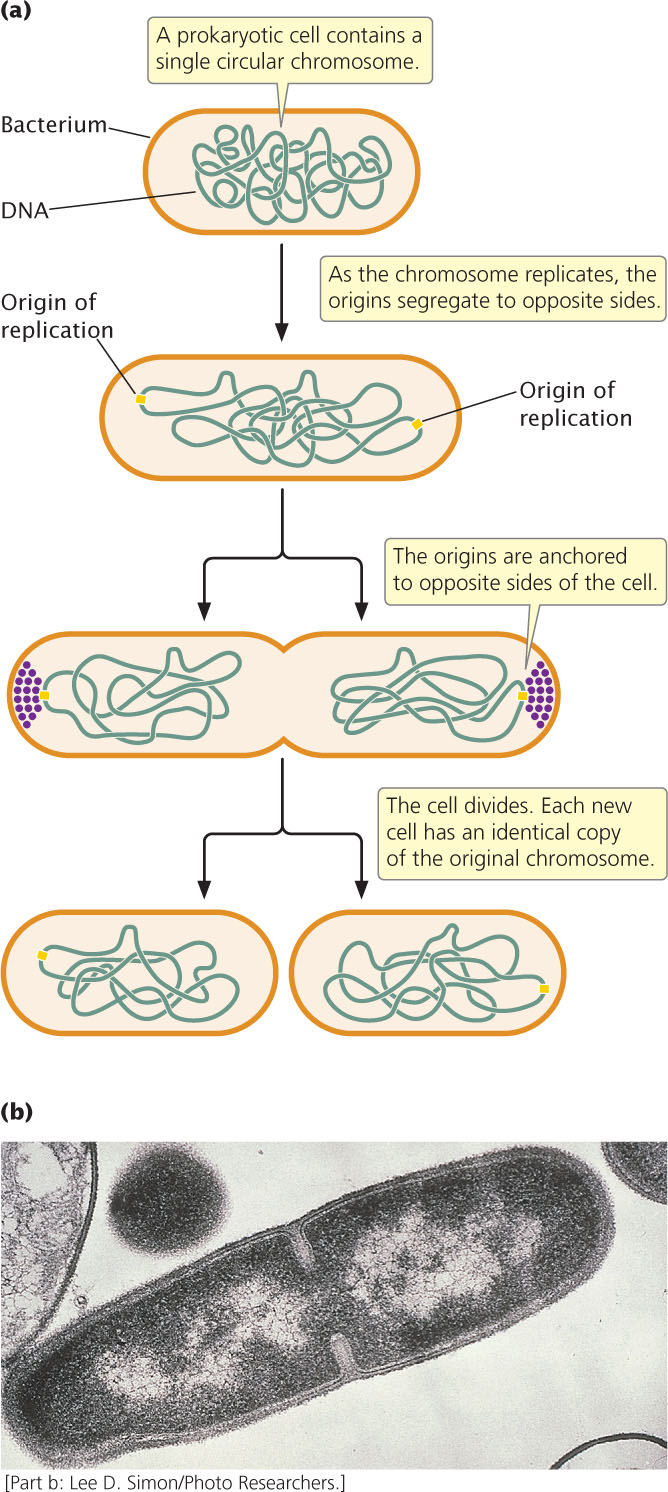
When prokaryotic cells reproduce, the circular chromosome of the bacterium replicates and the cell divides in a process called binary fission (Figure 2.5). Replication usually begins at a specific place on the bacterial chromosome, called the origin of replication. In a process that is not fully understood, the origins of the two newly replicated chromosomes move away from each other and toward opposite ends of the cell. In at least some bacteria, proteins bind near the replication origins and anchor the new chromosomes to the plasma membrane at opposite ends of the cell. Finally, a new cell wall forms between the two chromosomes, producing two cells, each with an identical copy of the chromosome. Under optimal conditions, some bacterial cells divide every 20 minutes. At this rate, a single bacterial cell could produce a billion descendants in a mere 10 hours.
Eukaryotic Cell Reproduction
Like prokaryotic cell reproduction, eukaryotic cell reproduction requires the processes of DNA replication, copy separation, and division of the cytoplasm. However, the presence of multiple DNA molecules requires a more-complex mechanism to ensure that exactly one copy of each molecule ends up in each of the new cells.
Eukaryotic chromosomes are separated from the cytoplasm by the nuclear envelope. The nucleus has a highly organized internal scaffolding called the nuclear matrix. This matrix consists of a network of protein fibers that maintains precise spatial relations among the nuclear components and takes part in DNA replication, the expression of genes, and the modification of gene products before they leave the nucleus. We will now take a closer look at the structure of eukaryotic chromosomes.
21
Eukaryotic ChromosomesEach eukaryotic species has a characteristic number of chromosomes per cell: potatoes have 48 chromosomes, fruit flies have 8, and humans have 46. There appears to be no special relation between the complexity of an organism and its number of chromosomes per cell.
In most eukaryotic cells, there are two sets of chromosomes. The presence of two sets is a consequence of sexual reproduction: one set is inherited from the male parent and the other from the female parent. Each chromosome in one set has a corresponding chromosome in the other set, together constituting a homologous pair (Figure 2.6). Human cells, for example, have 46 chromosomes, constituting 23 homologous pairs.
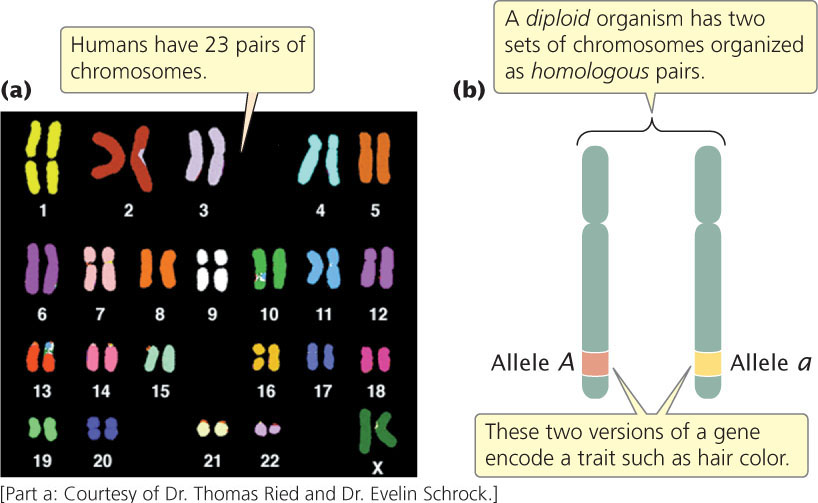
The two chromosomes of a homologous pair are usually alike in structure and size, and each carries genetic information for the same set of hereditary characteristics (the sex chromosomes are an exception and will be discussed in Chapter 4). For example, if a gene on a particular chromosome encodes a characteristic such as hair color, another copy of the gene (each copy is called an allele) at the same position on that chromosome’s homolog also encodes hair color. However, these two alleles need not be identical: one might encode brown hair and the other might encode blond hair. Cells that carry two sets of genetic information are diploid. In general, the ploidy of the cell indicates how many sets of genetic information the cell possesses. Reproductive cells (such as eggs, sperm, and spores) and even nonreproductive cells of some eukaryotic organisms may contain a single set of chromosomes and would be haploid. The cells of some organisms contain more than two sets of genetic information and are therefore called polyploid (see Chapter 8).
CONCEPTS
Cells reproduce by copying and separating their genetic information and then dividing. Because eukaryotes possess multiple chromosomes, mechanisms exist to ensure that each new cell receives one copy of each chromosome. Most eukaryotic cells are diploid, and their two chromosome sets can be arranged in homologous pairs. Haploid cells contain a single set of chromosomes.
 CONCEPT CHECK 2
CONCEPT CHECK 2
Diploid cells have
- two chromosomes
- two sets of chromosomes
- one set of chromosomes
- two pairs of homologous chromosomes
Chromosome StructureThe chromosomes of eukaryotic cells are larger and more complex than those found in prokaryotes, but each unreplicated chromosome nevertheless consists of a single molecule of DNA. Although linear, the DNA molecules in eukaryotic chromosomes are highly folded and condensed; if stretched out, some human chromosomes would be several centimeters long—thousands of times as long as the span of a typical nucleus. To package such a tremendous length of DNA into this small volume, each DNA molecule is coiled around histone proteins and tightly packed, forming a rod-shaped chromosome. Most of the time, the chromosomes are thin and difficult to observe, but before cell division, they condense further into thick, readily observed structures; it is at this stage that chromosomes are usually studied.
22
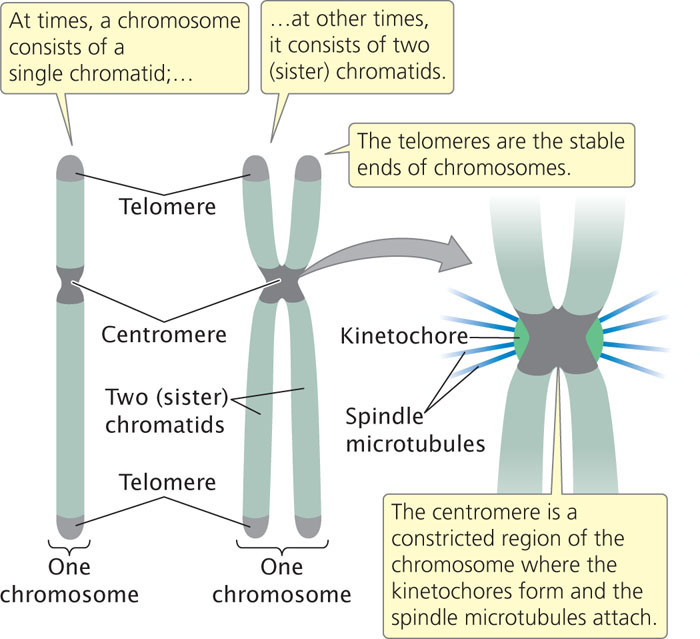
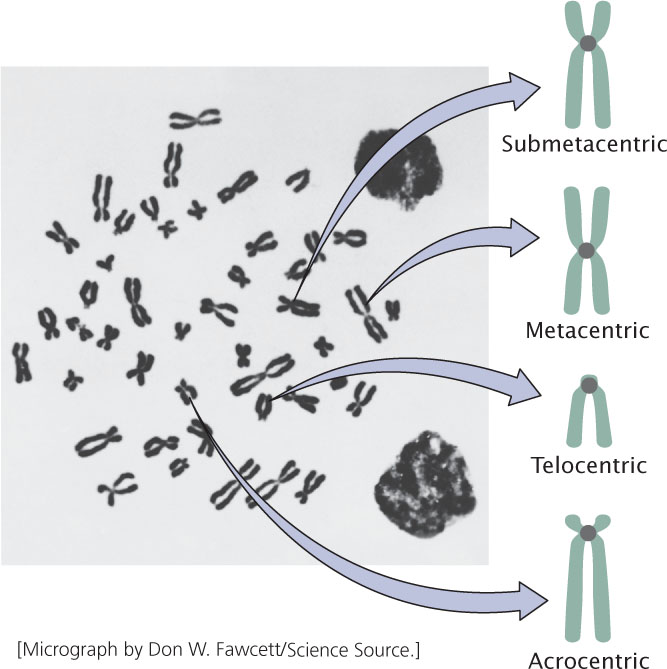
A functional chromosome has three essential elements: a centromere, a pair of telomeres, and origins of replication. The centromere is the attachment point for spindle microtubules—the filaments responsible for moving chromosomes in cell division (Figure 2.7). The centromere appears as a constricted region. Before cell division, a multiprotein complex called the kinetochore assembles on the centromere; later, spindle microtubules attach to the kinetochore. Chromosomes lacking a centromere cannot be drawn into the newly formed nuclei; these chromosomes are lost, often with catastrophic consequences for the cell. On the basis of the location of the centromere, chromosomes are classified into four types: metacentric, submetacentric, acrocentric, and telocentric (Figure 2.8).
Telomeres are the natural ends, the tips, of a whole linear chromosome (see Figure 2.7). Just as plastic tips protect the ends of a shoelace, telomeres protect and stabilize the chromosome ends. If a chromosome breaks, producing new ends, the chromosome is degraded at the newly broken ends. Telomeres provide chromosome stability. Research shows that telomeres also participate in limiting cell division and may play important roles in aging and cancer (discussed in Chapter 12).
23
Origins of replication are the sites where DNA synthesis begins; they are not easily observed by microscopy. Their structure and function will be discussed in more detail in Chapter 12. In preparation for cell division, each chromosome replicates, making a copy of itself, as already mentioned. These two initially identical copies, called sister chromatids, are held together at the centromere (see Figure 2.7). Each sister chromatid consists of a single molecule of DNA.
CONCEPTS
Sister chromatids are copies of a chromosome held together at the centromere. Functional chromosomes contain centromeres, telomeres, and origins of replication. The kinetochore is the point of attachment for the spindle microtubules; telomeres are the stabilizing ends of a chromosome; origins of replication are sites where DNA synthesis begins.
CONCEPT CHECK 3
What would be the result if a chromosome did not have a kinetochore?
The Cell Cycle and Mitosis
The cell cycle is the life story of a cell, the stages through which it passes from one division to the next (Figure 2.9). This process is critical to genetics because, through the cell cycle, the genetic instructions for all characteristics are passed from parent to daughter cells. A new cycle begins after a cell has divided and produced two new cells. Each new cell metabolizes, grows, and develops. At the end of its cycle, the cell divides to produce two cells, which can then undergo additional cell cycles. Progression through the cell cycle is regulated at key transition points called checkpoints, which allow or prohibit the cell’s progression to the next stage. Checkpoints ensure that all cellular components are present and in good working order, and are necessary to prevent cells with damaged or missing chromosomes from proliferating. Defects in checkpoints can lead to unregulated cell growth, as is seen in some cancers. The molecular basis of these checkpoints will be discussed in Chapter 23.
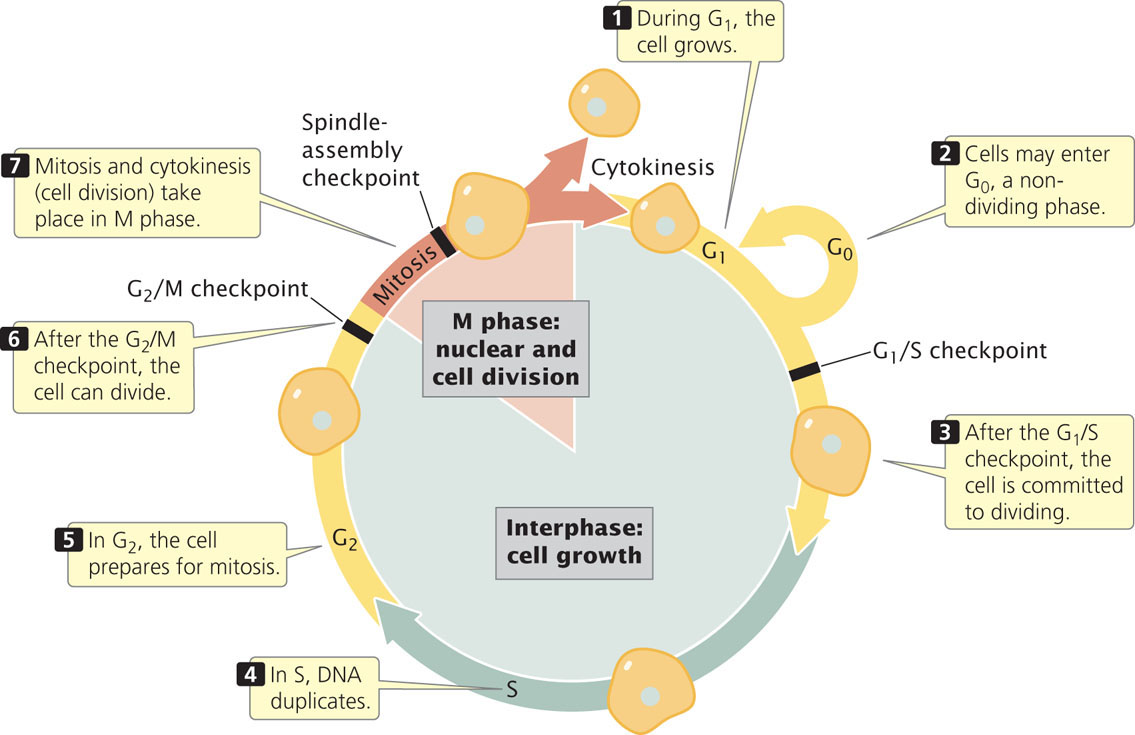
The cell cycle consists of two major phases. The first is interphase, the period between cell divisions, in which the cell grows, develops, and functions. In interphase, critical events necessary for cell division also take place. The second major phase is the M (mitotic) phase, the period of active cell division. The M phase includes mitosis, the process of nuclear division, and cytokinesis, or cytoplasmic division. Let’s take a closer look at the details of interphase and the M phase.
InterphaseInterphase is the extended period of growth and development between cell divisions. Although little activity can be observed with a light microscope, the cell is quite busy: DNA is being synthesized, RNA and proteins are being produced, and hundreds of biochemical reactions necessary for cellular functions are taking place. In addition to growth and development, interphase includes several checkpoints.
By convention, interphase is divided into three subphases: G1, S, and G2 (see Figure 2.9). Interphase begins with G1 (for gap 1). In G1, the cell grows, and proteins necessary for cell division are synthesized; this phase typically lasts several hours. Near the end of G1, a critical point, termed the G1/S checkpoint, holds the cell in G1 until the cell has all of the enzymes necessary for the replication of DNA. After this checkpoint has been passed, the cell is committed to divide.
24
Before reaching the G1/S checkpoint, cells may exit from the active cell cycle in response to regulatory signals and pass into a nondividing phase called G0, which is a stable state during which cells usually maintain a constant size. They can remain in G0 for an extended length of time, even indefinitely, or they can reenter G1 and the active cell cycle. Many cells never enter G0; rather, they cycle continuously.
After G1, the cell enters the S phase (for DNA synthesis), in which each chromosome duplicates. Although the cell is committed to divide after the G1/S checkpoint has been passed, DNA synthesis must take place before the cell can proceed to mitosis. If DNA synthesis is blocked (by drugs or by a mutation), the cell will not be able to undergo mitosis. Before the S phase, each chromosome is unreplicated; after the S phase, each chromosome is composed of two chromatids (see Figure 2.7).
After the S phase, the cell enters G2 (gap 2). In this phase, several additional biochemical events necessary for cell division take place. The important G2/M checkpoint is reached near the end of G2. This checkpoint is passed only if the cell’s DNA is completely replicated and undamaged. Unreplicated or damaged DNA can inhibit the activation of some proteins that are necessary for mitosis to take place. After the G2/M checkpoint has been passed, the cell is ready to divide and enters the M phase. Although the length of interphase varies from cell type to cell type, a typical dividing mammalian cell spends about 10 hours in G1, 9 hours in S, and 4 hours in G2 (see Figure 2.9).
Throughout interphase, the chromosomes are in a relaxed, but by no means uncoiled state, and individual chromosomes cannot be seen with a microscope. This condition changes dramatically when interphase draws to a close and the cell enters the M (mitotic) phase.
M PhaseThe M phase is the part of the cell cycle in which the copies of the cell’s chromosomes (sister chromatids) separate and the cell undergoes division. The separation of sister chromatids in the M phase is a critical process that results in a complete set of genetic information for each of the resulting cells. Biologists usually divide the M phase into six stages: the five stages of mitosis (prophase, prometaphase, metaphase, anaphase, and telophase) illustrated in Figure 2.10, and cytokinesis. It’s important to keep in mind that the M phase is a continuous process and that its separation into these six stages is somewhat arbitrary.
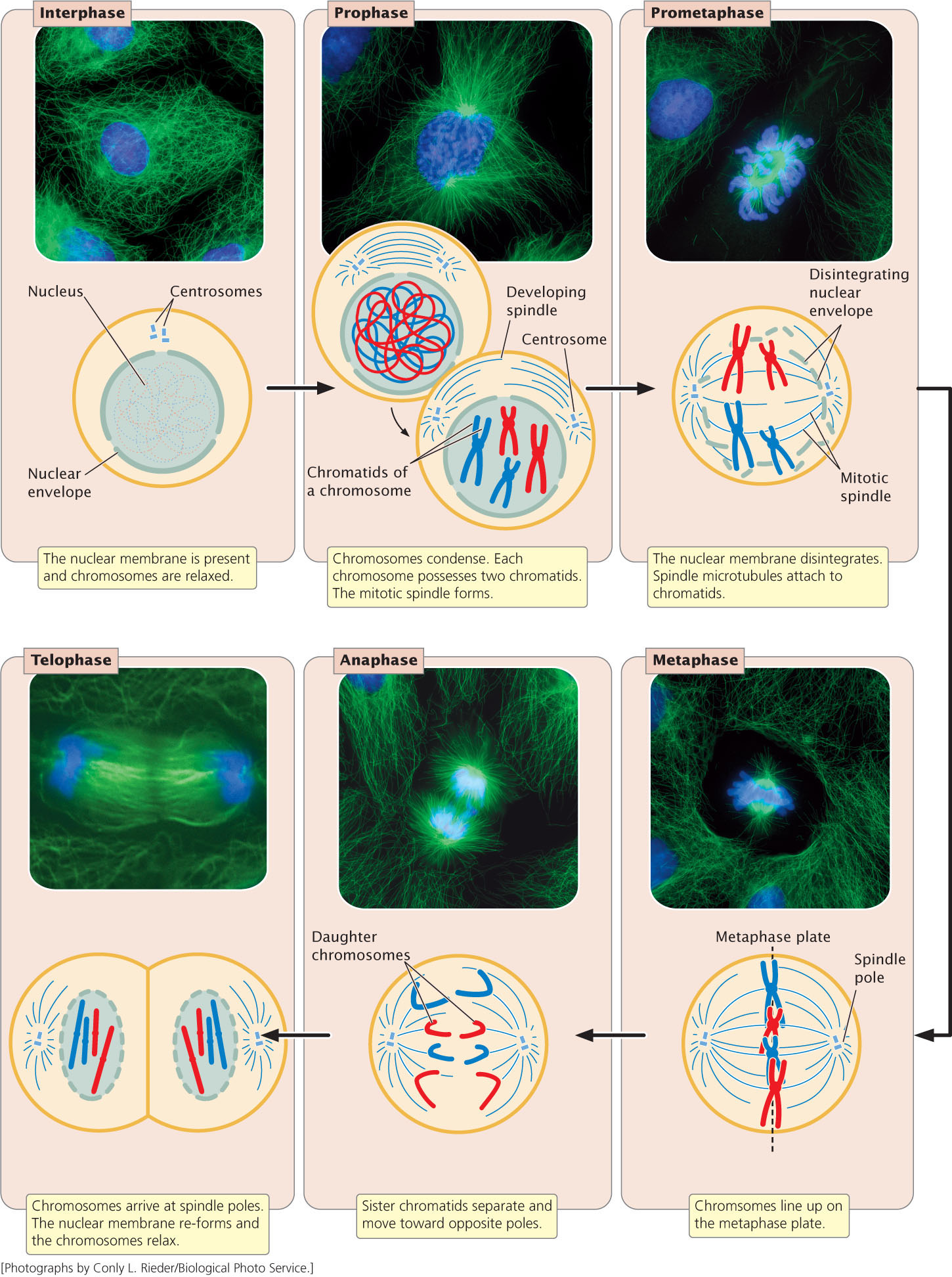
Prophase. As a cell enters prophase, the chromosomes become visible under a light microscope. Because the chromosome was duplicated in the preceding S phase, each chromosome possesses two chromatids attached at the centromere. The mitotic spindle, an organized array of microtubules that move the chromosomes in mitosis, forms. In animal cells, the spindle grows out from a pair of centrosomes that migrate to opposite sides of the cell. Within each centrosome is a special organelle, the centriole, which also is composed of microtubules. Some plant cells do not have centrosomes or centrioles, but they do have mitotic spindles.
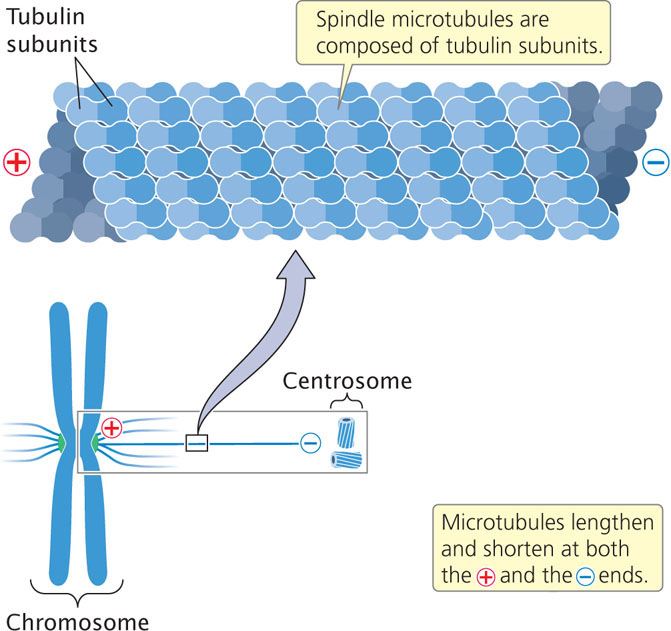
Prometaphase. Disintegration of the nuclear membrane marks the start of prometaphase. Spindle microtubules, which until now have been outside the nucleus, enter the nuclear region. The spindle microtubules are composed of subunits of a protein called tubulin (Figure 2.11). During prometaphase, tubulin molecules are added to and removed from the microtubules, causing them to undergo repeated cycles of growth and shrinkage. When the end of a microtubule encounters a kinetochore, the microtubule becomes stabilized. Eventually each chromosome becomes attached to microtubules from opposite spindle poles: for each chromosome, a microtubule from one of the centrosomes anchors to the kinetochore of one of the sister chromatids; a microtubule from the opposite centrosome then attaches to the other sister chromatid, anchoring the chromosome to both of the centrosomes. This arrangement is known as chromosome bi-orientation.
Metaphase. During metaphase, the chromosomes become arranged in a single plane, the metaphase plate, between the two centrosomes. The centrosomes, now at opposite ends of the cell with microtubules radiating outward and meeting in the middle of the cell, center at the spindle poles. A spindle-assembly checkpoint ensures that each chromosome is aligned on the metaphase plate and attached to spindle fibers from opposite poles.
The passage of a cell through the spindle-assembly checkpoint depends on tension generated at the kinetochore as the two conjoined chromatids are pulled in opposite directions by the spindle fibers. This tension is required for the cell to pass through the spindle-assembly checkpoint. If a microtubule attaches to one chromatid but not to the other, no tension is generated and the cell is unable to progress to the next stage of the cell cycle. The spindle-assembly checkpoint is able to detect even a single pair of chromosomes that are not properly attached to microtubules. The importance of this checkpoint is illustrated by cells that are defective in their spindle-assembly checkpoint; these cells often end up with abnormal numbers of chromosomes.
Anaphase. After the spindle-assembly checkpoint has been passed, the connection between sister chromatids breaks down and the sister chromatids separate. This chromatid separation marks the beginning of anaphase, during which the chromosomes move toward opposite spindle poles. Chromosome movement is due to the disassembly of tubulin molecules at both the kinetochore end (called the + end) and the spindle end (called the − end) of the spindle fiber (see Figure 2.11). Special proteins called molecular motors disassemble tubulin molecules from the spindle and generate forces that pull the chromosome toward the spindle pole.
25
26
Telophase. After the chromatids have separated, each is considered a separate chromosome. Telophase is marked by the arrival of the chromosomes at the spindle poles. The nuclear membrane re-forms around each set of chromosomes, producing two separate nuclei within the cell. The chromosomes relax and lengthen, once again disappearing from view. In many cells, division of the cytoplasm (cytokinesis) is simultaneous with telophase.
The major features of the cell cycle are summarized in Table 2.1. You can watch the cell cycle in motion by viewing Animation 2.1. This interactive animation allows you to determine what happens when different processes in the cycle fail.
Animation 2.1. This interactive animation allows you to determine what happens when different processes in the cycle fail. ![]() TRY PROBLEM 23
TRY PROBLEM 23
| Stage | Major Features |
|---|---|
| G0 phase | Stable, nondividing period of variable length. |
| Interphase | |
| G1 phase | Growth and development of the cell; G1/S checkpoint. |
| S phase | Synthesis of DNA. |
| G2 phase | preparation for division; G2/M checkpoint. |
| M phase | |
| Prophase | Chromosomes condense and mitotic spindle forms. |
| prometaphase | Nuclear envelope disintegrates, and spindle microtubules anchor to kinetochores. |
| Metaphase | Chromosomes align on the metaphase plate; spindle-assembly checkpoint. |
| Anaphase | Sister chromatids separate, becoming individual chromosomes that migrate toward spindle poles. |
| Telophase | Chromosomes arrive at spindle poles, the nuclear envelope re-forms, and the condensed chromosomes relax. |
| Cytokinesis | Cytoplasm divides; cell wall forms in plant cells. |
Genetic Consequences of the Cell Cycle
What are the genetically important results of the cell cycle? From a single cell, the cell cycle produces two cells that contain the same genetic instructions. The resulting daughter cells are genetically identical with each other and with their parent cell because DNA synthesis in the S phase creates an exact copy of each DNA molecule, giving rise to two genetically identical sister chromatids. Mitosis then ensures that one of the two sister chromatids from each replicated chromosome passes into each new cell.
Another genetically important result of the cell cycle is that each of the cells produced contains a full complement of chromosomes: there is no net reduction or increase in chromosome number. Each cell also contains approximately half the cytoplasm and organelle content of the original parental cell, but no precise mechanism analogous to mitosis ensures that organelles are evenly divided. Consequently, not all cells resulting from the cell cycle are identical in their cytoplasmic content.
CONCEPTS
The active cell cycle phases are interphase and the M phase. Interphase consists of G1, S, and G2. In G1, the cell grows and prepares for cell division; in the S phase, DNA synthesis takes place; in G2, other biochemical events necessary for cell division take place. Some cells enter a quiescent phase called G0. The M phase includes mitosis and cytokinesis and is divided into prophase, prometaphase, metaphase, anaphase, and telophase. The cell cycle produces two genetically identical cells each of which possesses a full complement of chromosomes.
CONCEPT CHECK 4
Which is the correct order of stages in the cell cycle?
- G1, S, prophase, metaphase, anaphase
- S, G1, prophase, metaphase, anaphase
- prophase, S, G1, metaphase, anaphase
- S, G1, anaphase, prophase, metaphase
27
CONNECTING CONCEPTS: Counting Chromosomes and DNA Molecules
The relations among chromosomes, chromatids, and DNA molecules frequently cause confusion. At certain times, chromosomes are unreplicated; at other times, each possesses two chromatids (see Figure 2.7). Chromosomes sometimes consist of a single DNA molecule; at other times, they consist of two DNA molecules. How can we keep track of the number of these structures in the cell cycle?
There are two simple rules for counting chromosomes and DNA molecules: (1) to determine the number of chromosomes, count the number of functional centromeres; (2) to determine the number of DNA molecules, first determine if sister chromatids are present. If sister chromatids are present, the chromosome has replicated and the number of DNA molecules is twice the number of chromosomes. If sister chromatids are not present, the chromosome has not replicated and the number of DNA molecules is the same as the number of chromosomes.
Let’s examine a hypothetical cell as it passes through the cell cycle (Figure 2.12). At the beginning of G1, this diploid cell has two complete sets of chromosomes, for a total of four chromosomes. Each chromosome is unreplicated and consists of a single molecule of DNA, and so there are four DNA molecules in the cell during G1. In the S phase, each DNA molecule is copied. The two resulting DNA molecules combine with histones and other proteins to form sister chromatids. Although the amount of DNA doubles in the S phase, the number of chromosomes remains the same because the sister chromatids are tethered together and share a single functional centromere. At the end of the S phase, this cell still contains four chromosomes, each with two sister chromatids; so 4 × 2 = 8 DNA molecules are present.
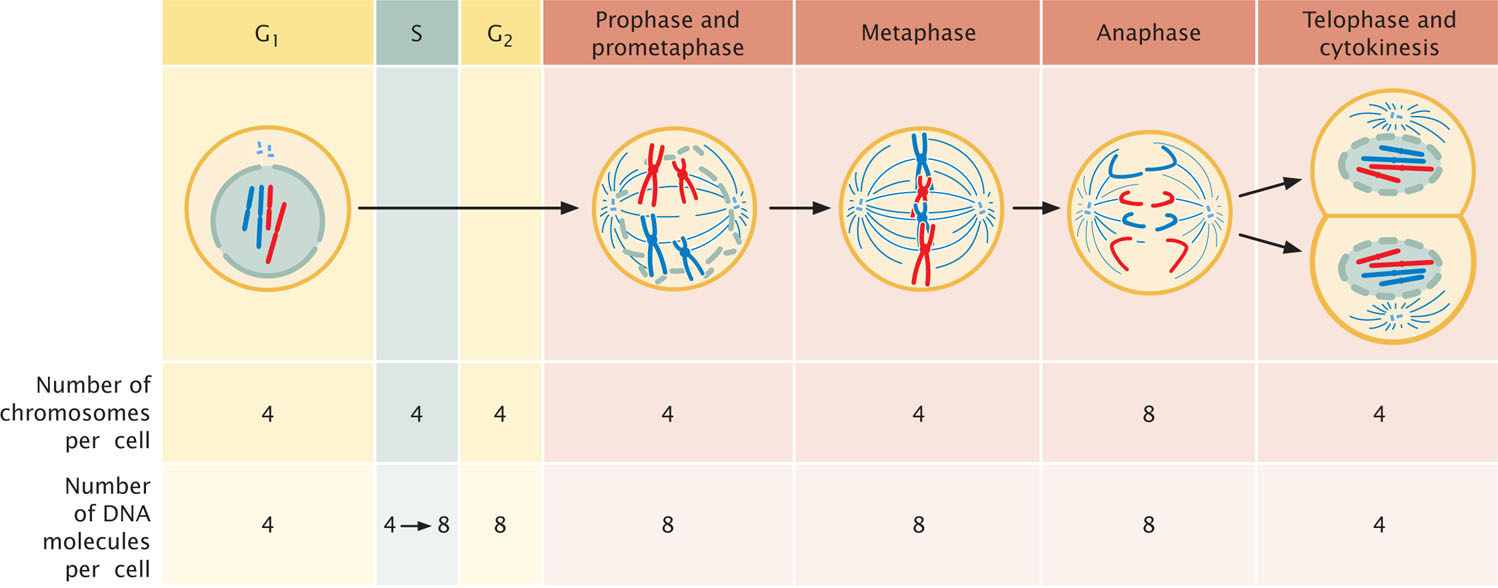
Through prophase, prometaphase, and metaphase, the cell has four chromosomes and eight DNA molecules. At anaphase, however, the sister chromatids separate. Each now has its own functional centromere, and so each is considered a separate chromosome. Until cytokinesis, the cell contains eight unreplicated chromosomes; thus, there are still eight DNA molecules present. After cytokinesis, the eight chromosomes (and eight DNA molecules) are distributed equally between two cells; so each new cell contains four chromosomes and four DNA molecules, the number present at the beginning of the cell cycle.
In summary, the number of chromosomes increases only in anaphase, when the two chromatids of a chromosome separate and become distinct chromosomes. The number of chromosomes decreases only through cytokinesis. The number of DNA molecules increases only in the S phase and decreases only through cytokinesis.![]() TRY PROBLEM 26
TRY PROBLEM 26