4.2 Sex-Linked Characteristics Are Determined by Genes on the Sex Chromosomes
In Chapter 3, we learned several basic principles of heredity that Mendel discovered from his crosses among pea plants. A major extension of these Mendelian principles is the pattern of inheritance exhibited by sex-linked characteristics, characteristics determined by genes located on the sex chromosomes. Genes on the X chromosome determine X-linked characteristics; those on the Y chromosome determine Y-linked characteristics. Because the Y chromosome of many organisms contains little genetic information, most sex-linked characteristics are X linked. Males and females differ in their sex chromosomes, so the pattern of inheritance for sex-linked characteristics differs from that exhibited by genes located on autosomal chromosomes.
X-Linked White Eyes in Drosophila
The first person to explain sex-linked inheritance was American biologist Thomas Hunt Morgan (Figure 4.11). Morgan began his career as an embryologist, but the discovery of Mendel’s principles inspired him to begin conducting genetic experiments, initially on mice and rats. In 1909, Morgan switched his research to Drosophila melanogaster; a year later, he discovered among the flies of his laboratory colony a single male that possessed white eyes, in stark contrast with the red eyes of normal fruit flies. This fly had a tremendous effect on Morgan’s career as a biologist and on the future of genetics.

To investigate the inheritance of the white-eyed characteristic in fruit flies, Morgan systematically carried out a series of genetic crosses. First, he crossed pure-breeding, red-eyed females with his white-eyed male, producing F1 progeny that had red eyes (Figure 4.12a). Morgan’s results from this initial cross were consistent with Mendel’s principles: a cross between a homozygous dominant individual and a homozygous recessive individual produces heterozygous offspring exhibiting the dominant trait. His results suggested that white eyes are a simple recessive trait. However, when Morgan crossed the F1 flies with one another, he found that all the female F2 flies possessed red eyes but that half the male F2 flies had red eyes and the other half had white eyes. This finding was clearly not the expected result for a simple recessive trait, which should appear in 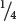 of both male and female F2 offspring.
of both male and female F2 offspring.
To explain this unexpected result, Morgan proposed that the locus affecting eye color is on the X chromosome (i.e., eye color is X linked). He recognized that the eye-color alleles are present only on the X chromosome; no homologous allele is present on the Y chromosome. Because the cells of females possess two X chromosomes, females can be homozygous or heterozygous for the eye-color alleles. The cells of males, on the other hand, possess only a single X chromosome and can carry only a single eye-color allele. Males therefore cannot be either homozygous or heterozygous but are said to be hemizygous for X-linked loci.
86
To verify his hypothesis that the white-eye trait is X linked, Morgan conducted additional crosses. He predicted that a cross between a white-eyed female and a red-eyed male would produce all red-eyed females and all white-eyed males (Figure 4.12b). When Morgan performed this cross, the results were exactly as predicted. Note that this cross is the reciprocal of the original cross and that the two reciprocal crosses produced different results in the F1 and F2 generations. Morgan also crossed the F1 heterozygous females with their white-eyed father, the red-eyed F2 females with white-eyed males, and white-eyed females with white-eyed males. In all of these crosses, the results were consistent with Morgan’s conclusion that the white-eye trait is an X-linked characteristic. You can view the results of Morgan’s crosses in  Animation 4.1.
Animation 4.1.
Nondisjunction and the Chromosome Theory of Inheritance
When Morgan crossed his original white-eyed male with homozygous red-eyed females, all 1237 of the progeny had red eyes, except for 3 white-eyed males. Morgan attributed these white-eyed F1 males to the occurrence of further random mutations. However, flies with these unexpected phenotypes continued to appear in his crosses. Although uncommon, they appeared far too often to be due to spontaneous mutation. Calvin Bridges, who was one of Morgan’s students, set out to investigate the genetic basis of these exceptions.
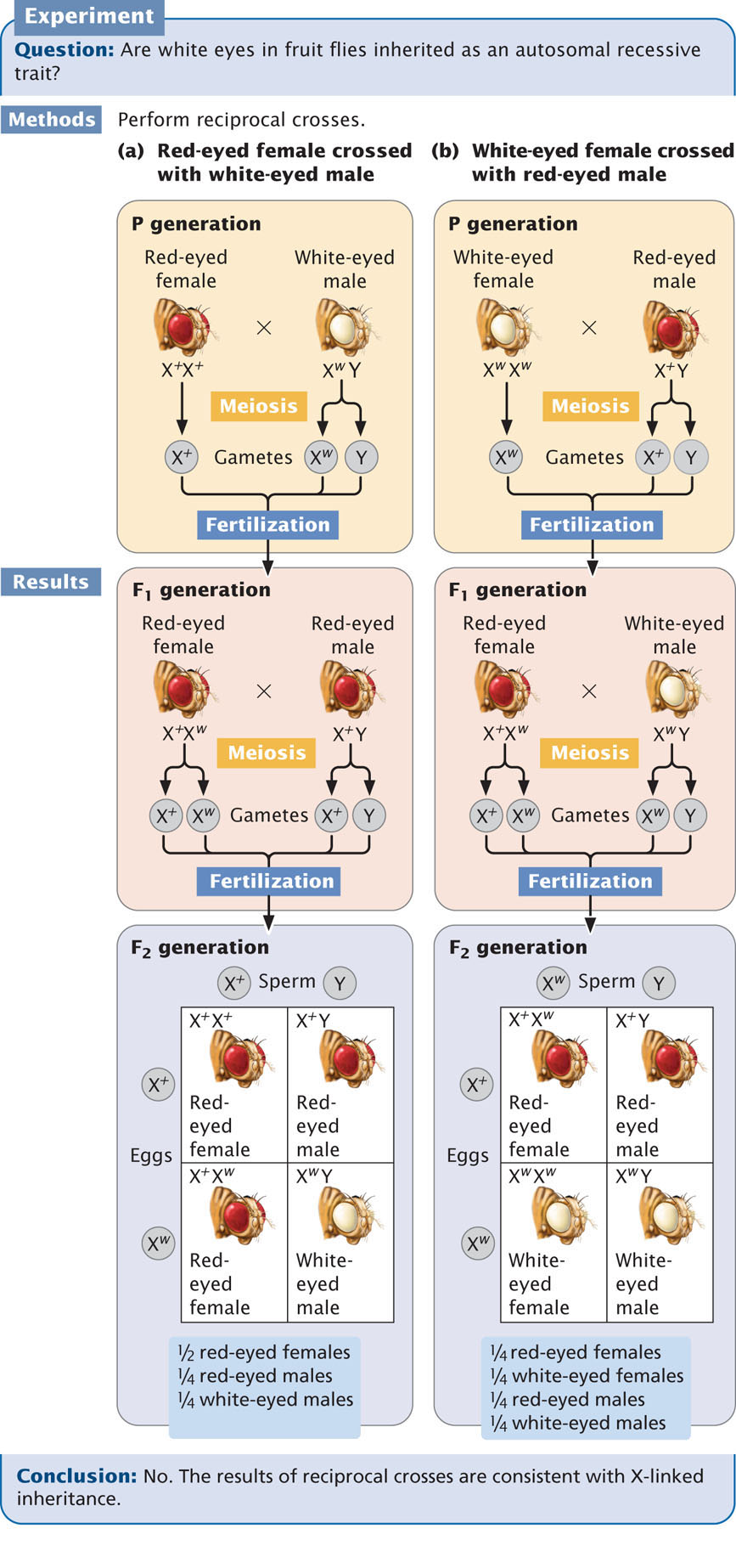
Bridges found that the exceptions arose only in certain strains of white-eyed flies. When he crossed one of these exceptional white-eyed females with a red-eyed male, about 5% of the male offspring had red eyes and about 5% of the female offspring had white eyes. In this cross, the expected result is that every male fly should inherit its mother’s X chromosome and should have the genotype Xw Y and white eyes (see the F1 progeny in Figure 4.12b). Every female fly should inherit a dominant red-eye allele on its father’s X chromosome, along with a white-eye allele on its mother’s X chromosome; thus, all the female progeny should be X+ Xw and have red eyes (see F1 progeny in Figure 4.12b). The continual appearance of red-eyed males and white-eyed females in this cross was therefore unexpected.
Bridges’s Explanation
To explain the appearance of red-eyed males and white-eyed females in his cross, Bridges hypothesized that the exceptional white-eyed females of this strain actually possessed two X chromosomes and a Y chromosome (Xw Xw Y). In Drosophila, flies with XXY sex chromosomes normally develop as females, in spite of possessing a Y chromosome (see Table 4.2). About 90% of the time, the two X chromosomes of the Xw Xw Y females separate from each other in anaphase I of meiosis, with an X and a Y chromosome entering one gamete and a single X entering another gamete (Figure 4.13). When these gametes are fertilized by sperm from a normal red-eyed male, white-eyed males and red-eyed females are produced. About 10% of the time, the two X chromosomes in the females fail to separate in anaphase I of meiosis, a phenomenon known as nondisjunction. When nondisjunction of the Xs occurs, half of the eggs receive two copies of the X chromosome and the other half receive only a Y chromosome (see Figure 4.13). When these eggs are fertilized by sperm from a normal red-eyed male, four combinations of sex chromosomes are produced. An egg with two X chromosomes that is fertilized by an X-bearing sperm produces an X+ XwXw zygote, which usually dies. When an egg carrying two X chromosomes is fertilized by a Y-bearing sperm, the resulting zygote is Xw Xw Y, which develops into a white-eyed female. An egg with only a Y chromosome that is fertilized by an X-bearing sperm produces an X+Y zygote, which develops into a normal red-eyed male. If the egg with only a Y chromosome is fertilized by a Y-bearing sperm, the resulting zygote has two Y chromosomes and no X chromosome and dies. Nondisjunction of the X chromosomes among Xw Xw Y white-eyed females therefore produces a few white-eyed females and red-eyed males, which is exactly what Bridges found in his crosses. The results of Bridges’s crosses are further explained in Animation 4.1.
87
Confirmation of Bridges’s Hypothesis
Bridges’s hypothesis predicted that the white-eyed females from these crosses would possess two X chromosomes and one Y chromosome and that the red-eyed males would possess a single X chromosome. To verify his hypothesis, Bridges examined the chromosomes of his flies and found precisely what he had predicted. The significance of Bridges’s study is not that it explained the appearance of an occasional odd fly in his culture but that he was able link the inheritance of a specific gene (w) to the presence of a specific chromosome (X). This association between genotype and chromosomes gave unequivocal evidence that sex-linked genes are located on the X chromosome and confirmed the chromosome theory of inheritance. ![]() TRY PROBLEM 22
TRY PROBLEM 22
88
CONCEPTS
By showing that the appearance of rare phenotypes is associated with the inheritance of particular chromosomes, Bridges proved that sex-linked genes are located on the X chromosome and that the chromosome theory of inheritance is correct.
 CONCEPT CHECK 5
CONCEPT CHECK 5
What was the genotype of the few F1 red-eyed males obtained by Bridges when he crossed a white-eyed female with a red-eyed male?
- X+
- XwX+Y
- X+Y
- X+X+Y
X-Linked Color Blindness in Humans
To further examine X-linked inheritance, let’s consider another X-linked characteristic: red–green color blindness in humans. The human eye perceives color through light-sensing cone cells that line the retina. Each cone cell contains one of three pigments capable of absorbing light of a particular wavelength; one absorbs blue light, a second absorbs red light, and a third absorbs green light. The human eye actually detects only three colors—red, green, and blue—but the brain mixes the signals from different cone cells to create the wide spectrum of colors that we perceive. Each of the three pigments is encoded by a separate locus; the locus for the blue pigment is found on chromosome 7, and those for the green and the red pigments lie close together on the X chromosome.
The most common types of human color blindness are caused by defects of the red and green pigments; we will refer to these conditions as red–green color blindness. Mutations that produce defective color vision are generally recessive and, because the genes encoding the red and the green pigments are located on the X chromosome, red–green color blindness is inherited as an X-linked recessive trait.
We will use the symbol Xc to represent an allele for red–green color blindness and the symbol X+ to represent an allele for normal color vision. Females possess two X chromosomes, so there are three possible genotypes among females: X+ X+ and X+ Xc, which produce normal vision, and Xc Xc, which produces color blindness. Males have only a single X chromosome and two possible genotypes: X+ Y, which produces normal vision, and Xc Y which produces color blindness.
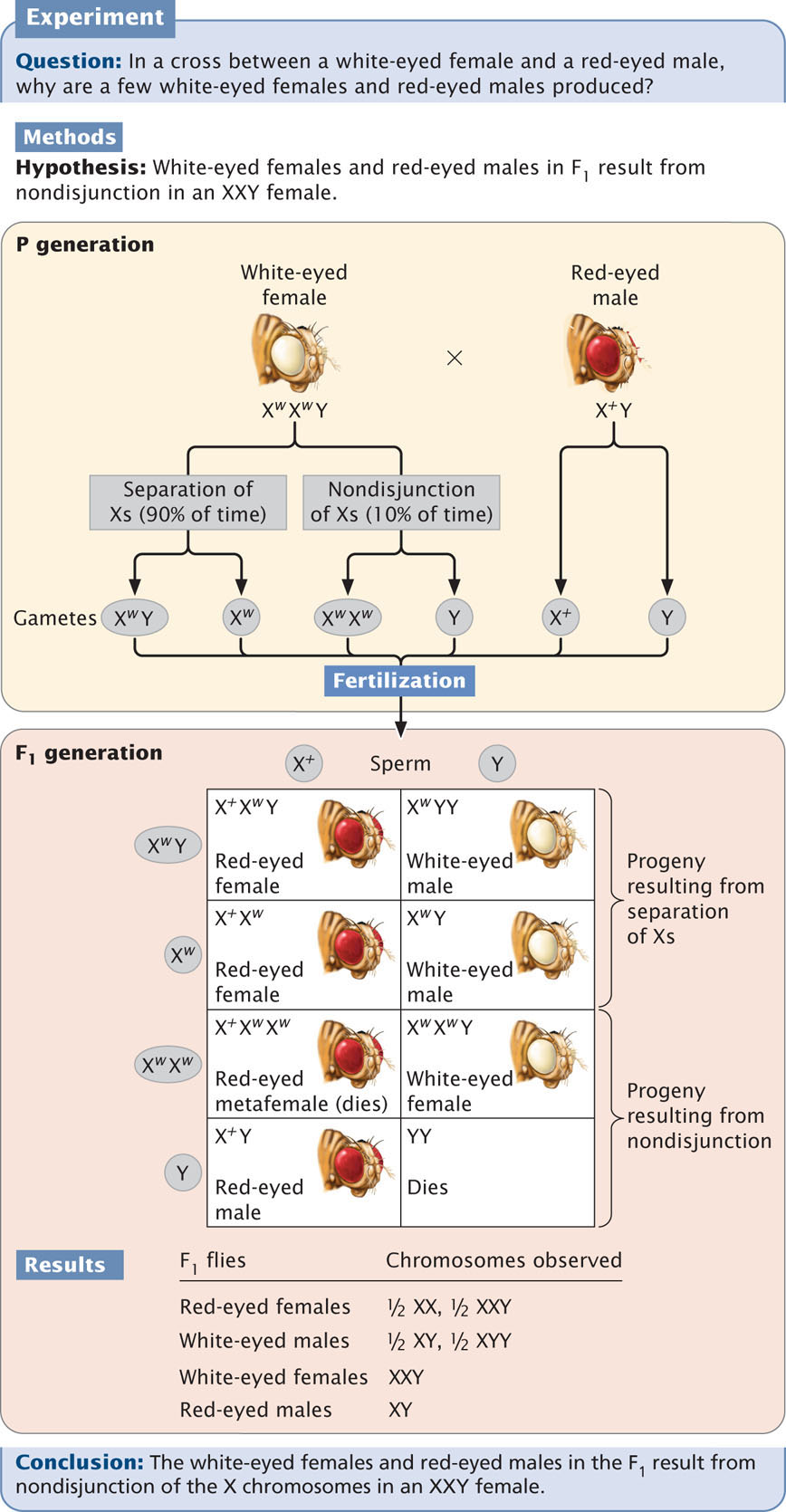
If a woman homozygous for normal color vision mates with a color-blind man (Figure 4.14a), all of the gametes produced by the woman will contain an allele for normal color vision. Half of the man’s gametes will receive the X chromosome with the color-blind allele, and the other half will receive the Y chromosome, which carries no alleles affecting color vision. When an Xc-bearing sperm unites with the X+-bearing egg, a heterozygous female with normal vision (X+Xc) is produced. When a Y-bearing sperm unites with the X-bearing egg, a hemizygous male with normal vision (X+ Y) is produced.
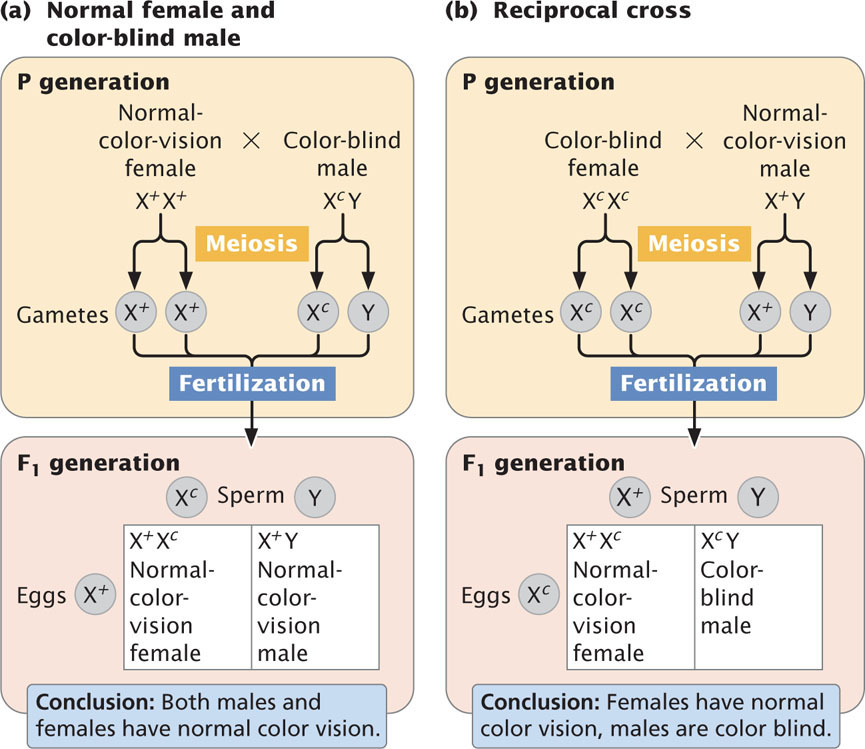
In the reciprocal cross between a color-blind woman and a man with normal color vision (Figure 4.14b), the woman produces only Xc-bearing gametes. The man produces some gametes that contain the X chromosome and others that contain the Y chromosome. Males inherit the X chromosome from their mothers and, because both of the mother’s X chromosomes bear the Xc allele, all the male offspring will be color blind. In contrast, females inherit an X chromosome from both parents; thus all the female offspring of this reciprocal cross will be heterozygous with normal vision. Females are color blind only when color-blind alleles have been inherited from both parents, whereas a colorblind male only needs to inherit a color-blind allele from his mother; for this reason, color blindness and most other rare X-linked recessive characteristics are more common in males.
In these crosses for color blindness, notice that an affected woman passes the X-linked recessive trait to her sons but not to her daughters, whereas an affected man passes the trait to his grandsons through his daughters but never to his sons. X-linked recessive characteristics may therefore appear to alternate between the sexes, appearing in females one generation and in males the next generation.
89
Recall that the X and Y chromosomes pair in meiosis because they are homologous at the small pseudoautosomal regions. Genes in these regions of the X and Y chromosome are homologous, just like those on autosomes, and they exhibit autosomal patterns of inheritance rather than the sex-linked inheritance seen for most genes on the X and Y chromosomes.
WORKED PROBLEMS
Now that we understand the pattern of X-linked inheritance, let’s apply our knowledge to answer a specific question.
Betty has normal vision, but her mother is color blind. Bill is color blind. If Bill and Betty marry and have a child together, what is the probability that the child will be color blind?
Solution
What information is required in your answer to the problem?
The probability that Bill and Betty’s child will be color blind.
What information is provided to solve the problem?
Phenotypes of Betty, Betty’s mother, and Bill.
Solution Steps
Because color blindness is an X-linked recessive trait, Betty’s color-blind mother must be homozygous for the color-blind allele (Xc Xc). Females inherit one X chromosome from each of their parents; so Betty must have inherited a color-blind allele from her mother. Because Betty has normal color vision, she must have inherited an allele for normal vision (X+) from her father; thus Betty is heterozygous (X+ Xc). Bill is color blind. Because males are hemizygous for X-linked alleles, he must be (Xc Y). A mating between Betty and Bill is represented as:
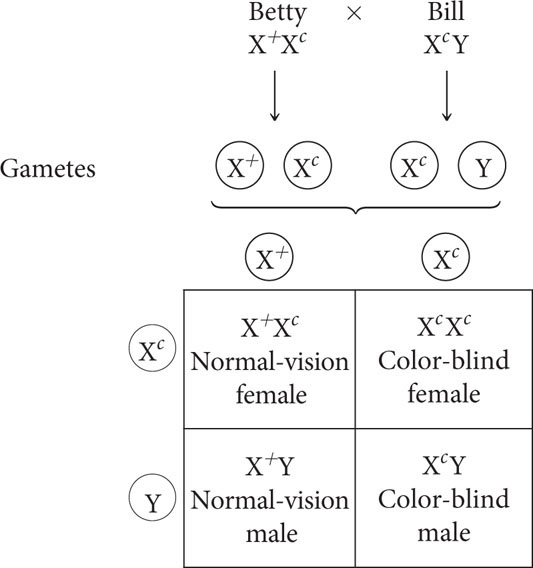
Thus, the overall probability that a child will be color blind is 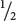 .
.
Get some additional practice with X-linked inheritance by working Problem 24 at the end of this chapter.
CONCEPTS
Characteristics determined by genes on the sex chromosomes are called sex-linked characteristics. Diploid females have two alleles at each X-linked locus, whereas diploid males possess a single allele at each X-linked locus. Females inherit X-linked alleles from both parents, but males inherit a single X-linked allele from their mothers.
CONCEPT CHECK 6
Hemophilia (reduced blood clotting) is an X-linked recessive disease in humans. A woman with hemophilia mates with a man who exhibits normal blood clotting. What is the probability that their child will have hemophilia?
Symbols for X-Linked Genes
There are several different ways to record genotypes for X-linked traits. Sometimes the genotypes are recorded in the same way as for autosomal characteristics. In this case, the hemizygous males are simply given a single allele: for example, the genotype of a female Drosophila with white eyes is ww, and the genotype of a white-eyed hemizygous male is w. Another method is to include the Y chromosome, designating it with a diagonal slash (/). With this method, the white-eyed female’s genotype is still ww and the white-eyed male’s genotype is w/. Perhaps the most useful method is to write the X and Y chromosomes in the genotype, designating the X-linked alleles with superscripts, as is done in this chapter. With this method, a white-eyed female is XwXw and a white-eyed male is XwY. The use of Xs and Ys in the genotype has the advantage of reminding us that the genes are X linked and that the male must always have a single allele, inherited from the mother.
Z-Linked Characteristics
In organisms with ZZ-ZW sex determination, the males are the homogametic sex (ZZ) and carry two sex-linked (usually referred to as Z-linked) alleles; thus males may be homozygous or heterozygous. Females are the heterogametic sex (ZW) and possess only a single Z-linked allele. The inheritance of Z-linked characteristics is the same as that of X-linked characteristics, except that the pattern of inheritance in males and females is reversed.
An example of a Z-linked characteristic is the cameo phenotype in Indian blue peafowl (Pavo eristatus). In these birds, the wild-type plumage is a glossy metallic blue. The female peafowl is ZW and the male is ZZ. Cameo plumage, which produces brown feathers, results from a Z-linked allele (Zca) that is recessive to the wild-type blue allele (ZCa+). If a blue-colored female (ZCa+W) is crossed with a cameo male (Zca Zca), all the F1 females are cameo (Zca W) and all the F1 males are blue (ZCa+Zca), as shown in Figure 4.15. When the F1 are interbred,
of the F2 are blue males (ZCa+Zca),
are blue females (ZCa+W),
are cameo males (Zca Zca), and
are cameo females (Zca W). The reciprocal cross of a cameo female with a homozygous blue male produces an F1 generation in which all offspring are blue and an F2 generation consisting of
blue males (ZCa+ZCa+ and ZCa+Zca),
blue females (ZCa+W), and
cameo females (Zca W).
90
In organisms with ZZ-ZW sex determination, the female always inherits her W chromosome from her mother, and she inherits her Z chromosome, along with any Z-linked alleles, from her father. In this system, the male inherits Z chromosomes, along with any Z-linked alleles, from both his mother and his father. This pattern of inheritance is the reverse of that of X-linked alleles in organisms with XX-XY sex determination. ![]() TRY PROBLEM 33
TRY PROBLEM 33
Y-Linked Characteristics
Y-linked traits—also called holandric traits—exhibit a distinct pattern of inheritance. These traits are present only in males, because only males possess a Y chromosome, and are always inherited from the father. Furthermore, all male offspring of a male with a Y-linked trait will display this trait, because every male inherits the Y chromosome from his father. ![]() TRY PROBLEM 45
TRY PROBLEM 45
Evolution of the Y Chromosome
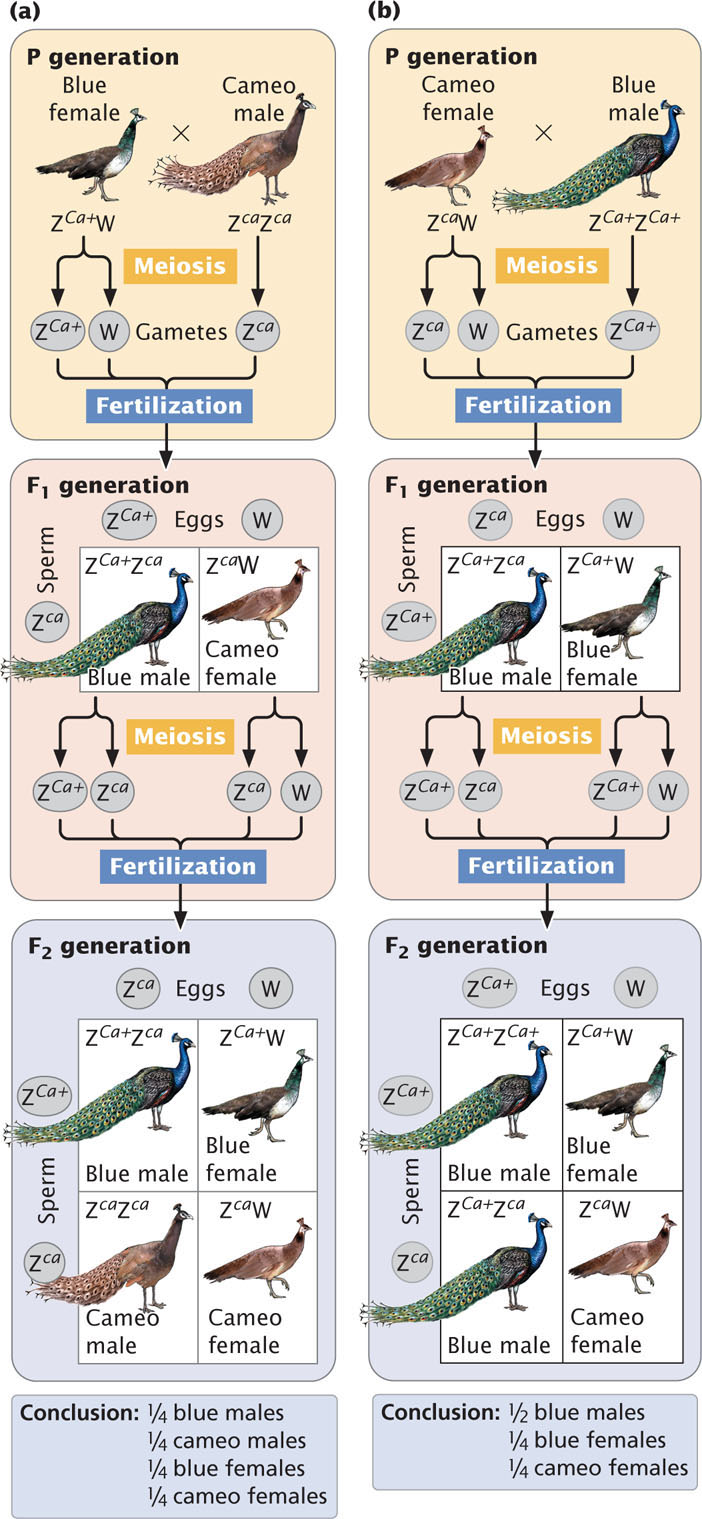
Research on sex chromosomes has led to the conclusion that the X and Y chromosomes in many organisms evolved from a pair of autosomes. The first step in this evolutionary process took place when one member of a pair of autosomes acquired a gene that determines maleness, such as the SRY gene found in humans today (Figure 4.16). This step took place in mammals about 250 million years ago. Any individual organism with a copy of the chromosome containing this gene then became male. Additional mutations occurred on the proto-Y chromosome affecting traits that are beneficial only in males, such as the bright coloration male birds use to attract females and the antlers a male elk uses in competition with other males. The genes that encode these types of traits are advantageous only if they are present in males. To prevent genes that encode male traits from appearing in females, crossing over was suppressed for most of the length of the X and Y chromosomes in meiosis. Crossing over can still take place between the two X chromosomes in females, but there is little crossing over between the X and the Y chromosomes, except for small pseudoautosomal regions in which the X and the Y chromosomes continue to pair and segregate in meiosis, as stated earlier.
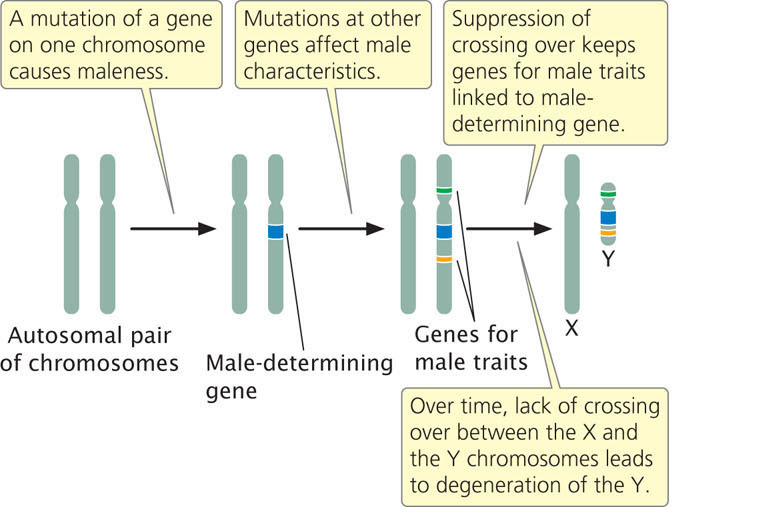
For reasons that are beyond the scope of the discussion here, the lack of crossing over led to (and continues to lead to) an accumulation of mutations and the loss of genetic material from the Y chromosome (see Figure 4.16). Over millions of years, the Y chromosome slowly degenerated, losing DNA and genes until it became greatly reduced in size and contained little genetic information. This degeneration produced the Y chromosome found in males today. Indeed, the Y chromosomes of humans and many other organisms are usually small and contain little genetic information; therefore, few characteristics exhibit Y-linked inheritance. Some researchers have predicted that the human Y chromosome will continue to lose genetic information in the future and will completely disappear from the species in about 10 million years, a disheartening prospect for those of us with a Y chromosome (and perhaps some of those with two Xs). However, research published in 2012 suggests that decay of the human Y chromosome has come to a halt and no genes have been lost in the last 6 million years. Internal recombination within the Y chromosome (see next section) may have aided in slowing down or preventing the complete decay of the human Y chromosome.
91
Characteristics of The Human Y Chromosome
The genetic sequence of most of the human Y chromosome has now been determined as part of the Human Genome Project (see Chapter 19). This work reveals that about two-thirds of the Y chromosome consists of short DNA sequences that are repeated many times and contain no active genes. The other third consists of just a few genes. Only about 350 genes have been identified on the human Y chromosome, compared with thousands on most chromosomes, and only about half of those identified on the Y chromosome encode proteins. The function of most Y-linked genes is poorly understood; many appear to influence male sexual development and fertility. Some are expressed throughout the body, but many are expressed predominately or exclusively in the testes. Although the Y chromosome has relatively few genes, recent research in Drosophila suggests that the Y chromosome carries genetic elements that affect the expression of numerous genes on autosomal and X chromosomes.
A surprising feature revealed by sequencing is the presence of eight massive palindromic sequences on the Y chromosome. A palindrome is defined as a word, such as “rotator,” or sentence that reads the same backward and forward. A palindromic sequence in DNA reads the same on both strands of the double helix, creating two nearly identical copies stretching out from a central point, such as:
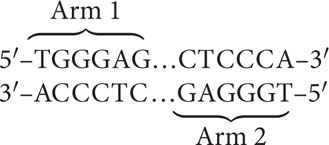
Thus, a palindromic sequence in DNA appears twice, very much like the two copies of a DNA sequence that are found on two homologous chromosomes. Indeed, recombination takes place between the two palindromic sequences on the Y chromosome. As already mentioned, the X and the Y chromosomes are not homologous at almost all of their sequences, and most of the Y chromosome does not undergo crossing over with the X chromosome. This lack of interchromosomal recombination leads to an accumulation of deleterious mutations on the Y chromosome and the loss of genetic material. Evidence suggests that the two arms of the Y chromosome recombine with each other, which partly compensates for the absence of recombination between the X and the Y chromosomes. This internal recombination may help to maintain some sequences and functions of genes on the Y chromosome and prevent its total degeneration.
Although the palindromes afford opportunities for recombination, which helps prevent the decay of the Y chromosome over evolutionary time, they occasionally have harmful effects. Recent research has revealed that recombination between the palindromes can lead to rearrangements of the Y chromosome that cause anomalies of sexual development. In some cases, recombination between the palindromes leads to deletion of the SRY gene, producing an XY female. In other cases, recombination deletes other Y-chromosome genes that take part in sperm production. Sometimes, recombination produces a Y chromosome with two centromeres, which may break as the centromeres are pulled in opposite directions in mitosis. The broken Y chromosomes may be lost in mitosis, resulting in XO cells and Turner syndrome.
CONCEPTS
Y-linked traits exhibit a distinct pattern of inheritance: they are present only in males, and all male offspring of a male with a Y-linked trait inherit the trait. Palindromic sequences within the Y chromosome can undergo internal recombination, but such recombination may lead to chromosome anomalies.
CONCEPT CHECK 7
What unusual feature of the Y chromosome allows some recombination among the genes found on it?
The use of Y-Linked Genetic Markers
DNA sequences in the Y chromosome undergo mutation with the passage of time and vary among individual males. These mutations create variations in DNA sequence—called genetic markers—that, like Y-linked traits, are passed from father to son and that can be used to study male ancestry. Although the markers themselves do not encode any physical traits, they can be detected with the use of molecular methods. Much of the Y chromosome is nonfunctional so mutations readily accumulate. Many of these mutations are unique; they arise only once and are passed down through the generations. Individual males possessing the same set of mutations are therefore related, and the distribution of these genetic markers on Y chromosomes provides clues about the genetic relationships of present-day people.
92
Y-linked markers have been used to study the offspring of Thomas Jefferson, principal author of the Declaration of Independence and third president of the United States. In 1802, a political enemy accused Jefferson of fathering a child by his slave Sally Hemings, but the evidence was circumstantial. Hemings, who worked in the Jefferson household and accompanied Jefferson on a trip to Paris, had five children. Jefferson was accused of fathering the first child, but rumors about the paternity of the other children circulated as well. Descendants of Hemings’s children maintained that they were descendants of the Jefferson line, but some Jefferson descendants refused to recognize their claim.
To resolve this long-standing controversy, geneticists examined markers from the Y chromosomes of male-line descendants of Hemings’s first son (Thomas Woodson), her last son (Eston Hemings), and a paternal uncle of Thomas Jefferson with whom Jefferson had Y chromosomes in common—descendants of Jefferson’s uncle were used because Jefferson himself had no verified male descendants. Geneticists determined that Jefferson possessed a rare and distinctive set of genetic markers on his Y chromosome. The same markers were also found on the Y chromosomes of the male-line descendants of Eston Hemings. The probability of such a match arising by chance is less than 1%. The markers were not found on the Y chromosomes of the descendants of Thomas Woodson. Together with the circumstantial historical evidence, these matching markers strongly suggest that Jefferson fathered Eston Hemings but not Thomas Woodson.
Y-chromosome sequences have also been used extensively to examine past patterns of human migration and the genetic relationships among different human populations.
CONNECTING CONCEPTS: Recognizing Sex-Linked Inheritance
What features should we look for to identify a trait as sex linked? A common misconception is that any genetic characteristic in which the phenotypes of males and females differ must be sex linked. In fact, the expression of many autosomal characteristics differs between males and females. The genes that encode these characteristics are the same in both sexes, but their expression is influenced by sex hormones. The different sex hormones of males and females cause the same genes to generate different phenotypes in males and females.
Another misconception is that any characteristic that is found more frequently in one sex is sex linked. A number of autosomal traits are expressed more commonly in one sex than in the other. These traits are said to be sex influenced. Some autosomal traits are expressed in only one sex; these traits are said to be sex limited. Both sex-influenced and sex-limited characteristics will be considered in more detail in Chapter 5.
Several features of sex-linked characteristics make them easy to recognize. Y-linked traits are found only in males, but this fact does not guarantee that a trait is Y linked, because some autosomal characteristics are expressed only in males. A Y-linked trait is unique, however, in that all the male offspring of an affected male will express the father’s phenotype, and a Y-linked trait can be inherited only from the father’s side of the family. Thus, a Y-linked trait can be inherited only from the paternal grandfather (the father’s father), never from the maternal grandfather (the mother’s father).
X-linked characteristics also exhibit a distinctive pattern of inheritance. X linkage is a possible explanation when reciprocal crosses give different results. If a characteristic is X-linked, a cross between an affected male and an unaffected female will not give the same results as a cross between an affected female and an unaffected male. For almost all autosomal characteristics, reciprocal crosses give the same result. We should not conclude, however, that, when the reciprocal crosses give different results, the characteristic is X linked. Other sex-associated forms of inheritance, considered in Chapter 5, also produce different results in reciprocal crosses. The key to recognizing X-linked inheritance is to remember that a male always inherits his X chromosome from his mother, not from his father. Thus, an X-linked characteristic is not passed directly from father to son; if a male clearly inherits a trait from his father—and the mother is not heterozygous—it cannot be X linked.