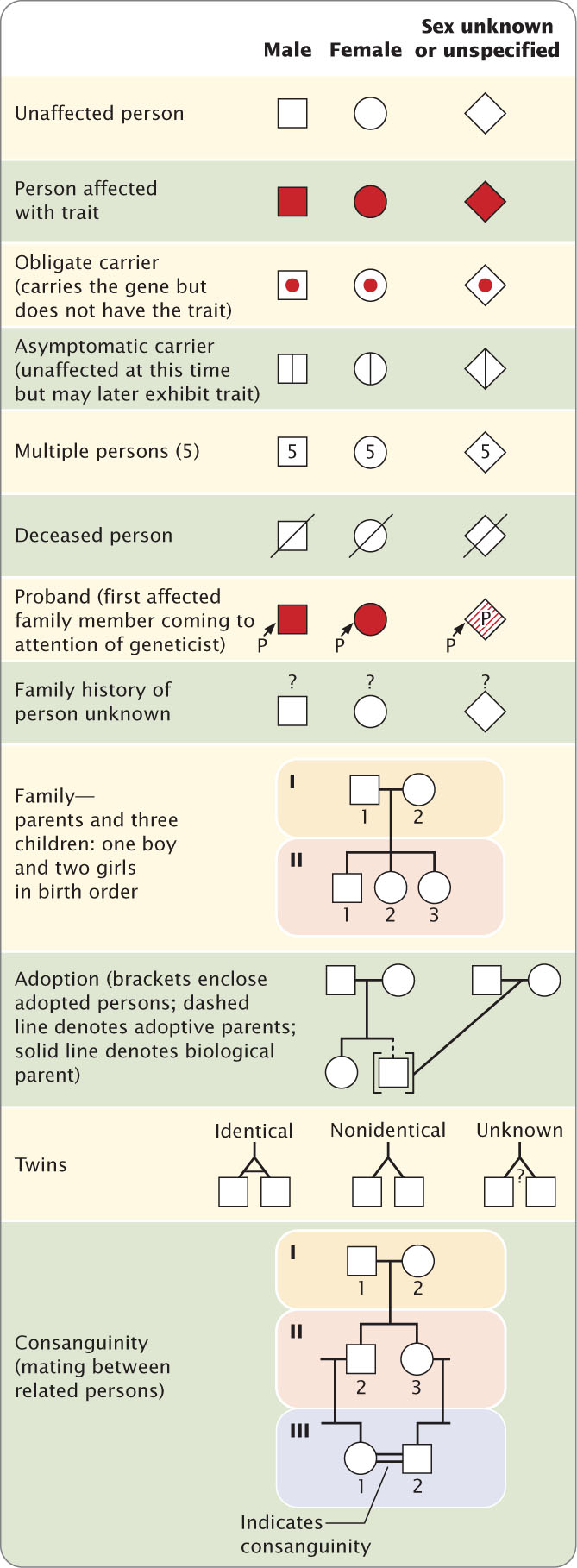
6.2 Geneticists Often Use Pedigrees to Study the Inheritance of Characteristics in Humans
An important technique used by geneticists to study human inheritance is the analysis of pedigrees. A pedigree is a pictorial representation of a family history, essentially a family tree that outlines the inheritance of one or more characteristics. When a particular characteristic or disease is observed in a person, a geneticist often studies the family of this affected person by drawing a pedigree.
Symbols Used in Pedigrees
The symbols commonly used in pedigrees are summarized in Figure 6.2. Males in a pedigree are represented by squares, females by circles. A horizontal line drawn between two symbols representing a man and a woman indicates a mating; children are connected to their parents by vertical lines extending downward from the parents. The pedigree shown in Figure 6.3a illustrates a family with Waardenburg syndrome, an autosomal dominant type of deafness that may be accompanied by fair skin, a white forelock, and visual problems (Figure 6.3b). Persons who exhibit the trait of interest are represented by filled circles and squares; in the pedigree of Figure 6.3a, the filled symbols represent members of the family who have Waardenburg syndrome. Unaffected members are represented by open circles and squares. The person from whom the pedigree is initiated is called the proband and is usually designated by an arrow (IV-2 in Figure 6.3a).
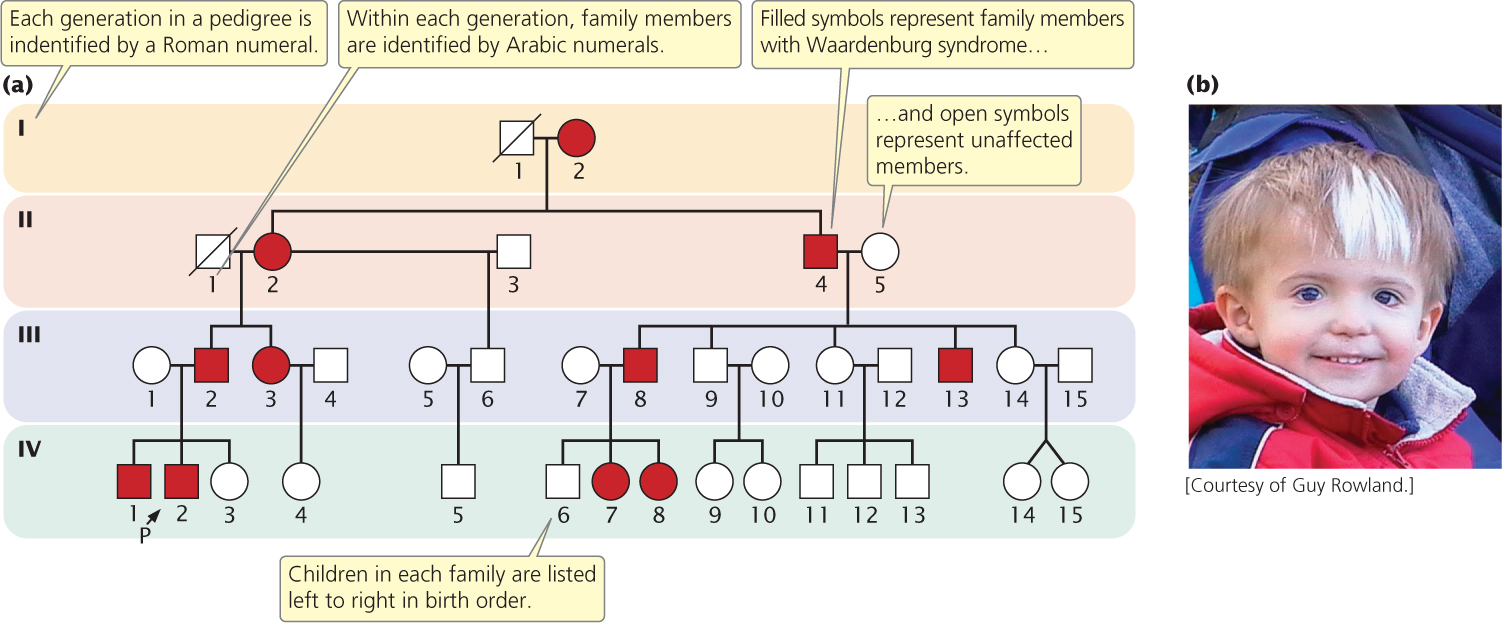
Let’s look closely at Figure 6.3 and consider some additional features of a pedigree. Each generation in a pedigree is identified by a Roman numeral; within each generation family members are assigned Arabic numerals, and children in each family are listed in birth order from left to right. Person II-4, a man with Waardenburg syndrome, mated with II-5, an unaffected woman, and they produced five children. The oldest of their children is III-8, a male with Waardenburg syndrome, and the youngest is III-14, an unaffected female. ![]() TRY PROBLEM 19a
TRY PROBLEM 19a
Analysis of Pedigrees
The limited number of offspring in most human families means that clear Mendelian ratios in a single pedigree are usually impossible to discern. Pedigree analysis requires a certain amount of genetic sleuthing, based on recognizing patterns associated with different modes of inheritance. For example, autosomal dominant traits should appear with equal frequency in both sexes and should not skip generations, provided that the trait is fully penetrant and not sex influenced.
Certain patterns may exclude the possibility of a particular mode of inheritance. For instance, a son inherits his X chromosome from his mother. If we observe that a trait is passed from father to son, we can exclude the possibility of X-linked inheritance. In the following sections, the traits discussed are assumed to be fully penetrant and rare.
142
Autosomal Recessive Traits
Autosomal recessive traits normally appear with equal frequency in both sexes (unless penetrance differs in males and females) and appear only when a person inherits two alleles for the trait, one from each parent. If the trait is uncommon, most parents of affected offspring are heterozygous and unaffected; consequently, the trait seems to skip generations (Figure 6.4). Frequently, a recessive allele may be passed for a number of generations without the trait appearing in a pedigree. Whenever both parents are heterozygous, approximately one-fourth of the offspring are expected to express the trait, but this ratio will not be obvious unless the family is large. In the rare event that both parents are affected by an autosomal recessive trait, all the offspring will be affected.
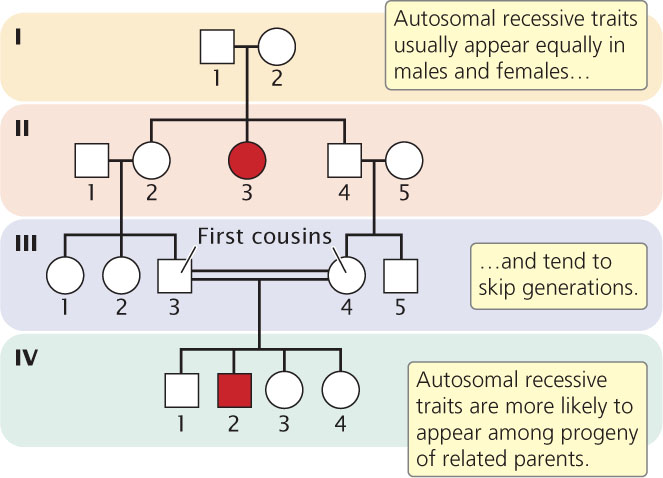
When a recessive trait is rare, persons from outside the family are usually homozygous for the normal allele. Thus, when an affected person mates with someone outside the family (aa × AA), usually none of the children will display the trait, although all will be carriers (i.e., heterozygous). A recessive trait is more likely to appear in a pedigree when two people within the same family mate, because there is a greater chance of both parents carrying the same recessive allele. Mating between closely related people is called consanguinity. In the pedigree shown in Figure 6.4, persons III-3 and III-4 are first cousins, and both are heterozygous for the recessive allele; when two heterozygotes mate, 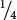 of their children are expected to have the recessive trait.
of their children are expected to have the recessive trait.
CONCEPTS
Autosomal recessive traits appear with equal frequency in males and females. Affected children are commonly born to unaffected parents who are carriers of the gene for the trait, and the trait tends to skip generations. Recessive traits appear more frequently among the offspring of consanguine matings.
 CONCEPT CHECK 1
CONCEPT CHECK 1
Autosomal recessive traits often appear in pedigrees in which there have been consanguine matings, because these traits
- tend to skip generations.
- appear only when both parents carry a copy of the gene for the trait, which is more likely when the parents are related.
- usually arise in children born to parents who are unaffected.
- appear equally in males and females.
143
A number of human metabolic diseases are inherited as autosomal recessive traits. One of them is Tay–Sachs disease. Children with Tay–Sachs disease appear healthy at birth but become listless and weak at about 6 months of age. Gradually, their physical and neurological conditions worsen, leading to blindness, deafness, and, eventually, death at 2 to 3 years of age. The disease results from the accumulation of a lipid called GM2 ganglioside in the brain. A normal component of brain cells, GM2 ganglioside is usually broken down by an enzyme called hexosaminidase A, but children with Tay–Sachs disease lack this enzyme. Excessive GM2 ganglioside accumulates in the brain, causing swelling and, ultimately, neurological symptoms. Heterozygotes have only one normal copy of the allele encoding hexosaminidase A and produce only about half the normal amount of the enzyme. However, this amount is enough to ensure that GM2 ganglioside is broken down normally, and heterozygotes are usually healthy.
Autosomal Dominant Traits
Autosomal dominant traits appear in both sexes with equal frequency, and both sexes are capable of transmitting these traits to their offspring. Every person with a dominant trait must have inherited the allele from at least one parent; autosomal dominant traits therefore do not skip generations (Figure 6.5). Exceptions to this rule arise when people acquire the trait as a result of a new mutation or when the trait has reduced penetrance.
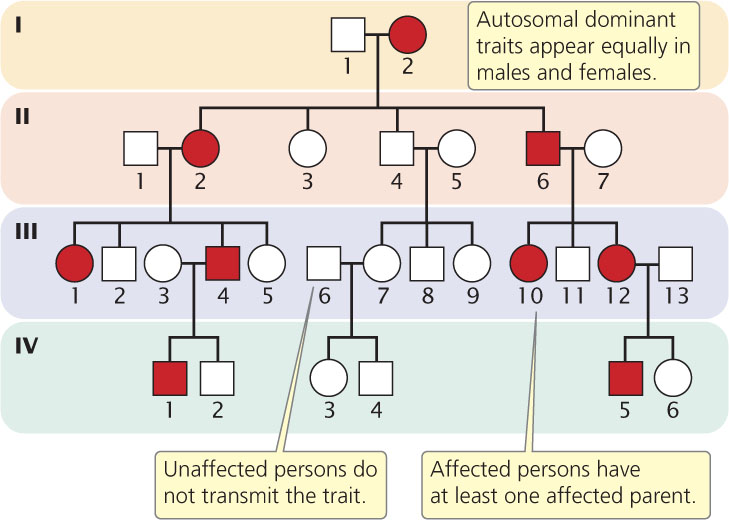
If an autosomal dominant allele is rare, most people displaying the trait are heterozygous. When one parent is affected and heterozygous and the other parent is unaffected, approximately 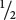 of the offspring will be affected. If both parents have the trait and are heterozygous, approximately
of the offspring will be affected. If both parents have the trait and are heterozygous, approximately 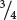 of the children will be affected. Unaffected people do not transmit the trait to their descendants, provided that the trait is fully penetrant. In Figure 6.5, we see that none of the descendants of II-4 (who is unaffected) have the trait.
of the children will be affected. Unaffected people do not transmit the trait to their descendants, provided that the trait is fully penetrant. In Figure 6.5, we see that none of the descendants of II-4 (who is unaffected) have the trait.
CONCEPTS
Autosomal dominant traits appear in both sexes with equal frequency. An affected person has an affected parent (unless the person carries new mutations), and the trait does not skip generations. Unaffected persons do not transmit the trait.
CONCEPT CHECK 2
When might you see an autosomal dominant trait skip generations?
A trait that is usually considered to be autosomal dominant is familial hypercholesterolemia, an inherited disease in which blood cholesterol is greatly elevated owing to a defect in cholesterol transport. Cholesterol is transported throughout the body in small soluble particles called lipoproteins (Figure 6.6). A principal lipoprotein in the transport of cholesterol is low-density lipoprotein (LDL). When an LDL molecule reaches a cell, it attaches to an LDL receptor, which then moves the LDL through the cell membrane into the cytoplasm, where it is broken down and its cholesterol is released for use by the cell.
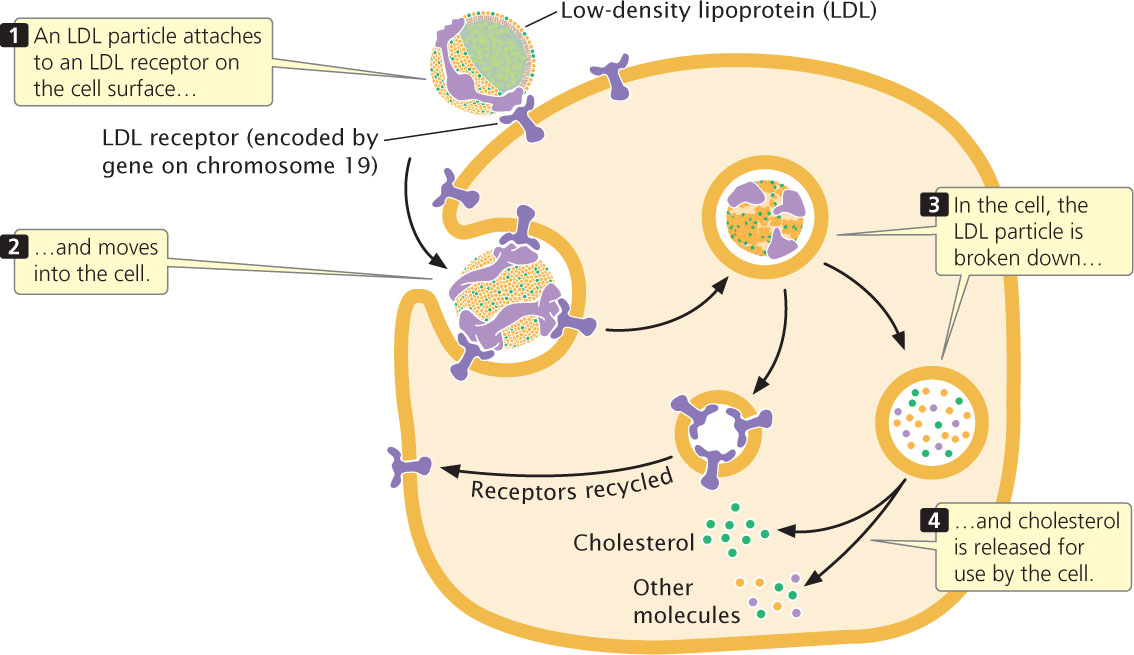
Familial hypercholesterolemia occurs when there is a defect in the gene that normally encodes the LDL receptor. The disease is usually considered an autosomal dominant disorder because heterozygotes are deficient in LDL receptors and have elevated blood levels of cholesterol, leading to increased risk of coronary artery disease. Persons heterozygous for familial hypercholesterolemia have blood LDL levels that are twice normal and usually have heart attacks by the age of 35.
Very rarely, a person inherits two defective alleles for the LDL receptor. Such persons don’t make any functional LDL receptors; their blood cholesterol levels are more than six times normal, and they may suffer a heart attack as early as age 2 and almost inevitably by age 20. Because homozygotes are more severely affected than heterozygotes, familial hypercholesterolemia is said to be incompletely dominant. However, homozygotes are rarely seen, and the common heterozygous form of the disease appears as a simple dominant trait in most pedigrees.
X-Linked Recessive Traits
X-linked recessive traits have a distinctive pattern of inheritance (Figure 6.7). First, these traits appear more frequently in males than in females because males need inherit only a single copy of the allele to display the trait, whereas females must inherit two copies of the allele, one from each parent, to be affected. Second, because a male inherits his X chromosome from his mother, affected males are usually born to unaffected mothers who carry an allele for the trait. Because the trait is passed from unaffected female to affected male to unaffected female, it tends to skip generations (see Figure 6.7). When a woman is heterozygous, approximately
of her sons will be affected and
of her daughters will be unaffected carriers. For example, we know that females I-2, II-2, and III-7 in Figure 6.7 are carriers because they transmit the trait to approximately half of their sons.
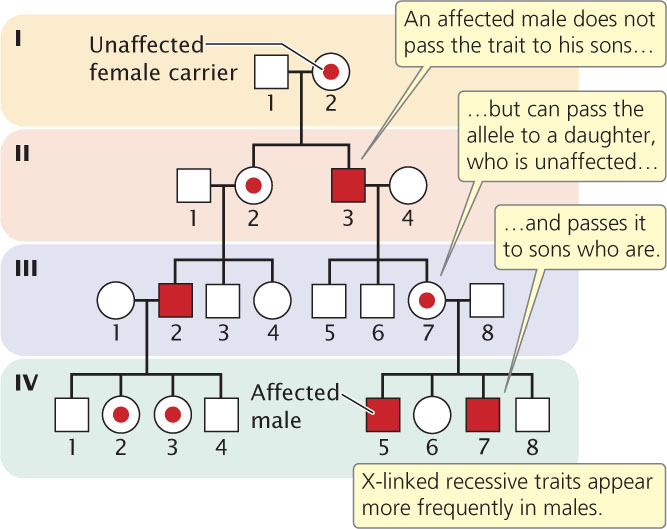
144
A third important characteristic of X-linked recessive traits is that they are not passed from father to son, because a son inherits his father’s Y chromosome, not his X. In Figure 6.7, there is no case in which both a father and his son are affected. All daughters of an affected man, however, will be carriers (if their mother is homozygous for the normal allele). When a woman displays an X-linked recessive trait, she must be homozygous for the trait, and all of her sons also will display the trait.
CONCEPTS
Rare X-linked recessive traits appear more often in males than in females and are not passed from father to son. Affected sons are usually born to unaffected mothers who are carriers of the gene for the trait; thus X-linked recessive traits tend to skip generations.
CONCEPT CHECK 3
How can you distinguish between an autosomal recessive trait with higher penetrance in males and an X-linked recessive trait?
An example of an X-linked recessive trait in humans is hemophilia A, also called classic hemophilia. Hemophilia results from the absence of a protein necessary for blood to clot. The complex process of blood clotting consists of a cascade of reactions that includes more than 13 different factors. For this reason, there are several types of clotting disorders, each due to a glitch in a different step of the clotting pathway.
Hemophilia A results from abnormal or missing factor VIII, one of the proteins in the clotting cascade. The gene for factor VIII is located on the tip of the long arm of the X chromosome; so hemophilia A is an X-linked recessive disorder. People with hemophilia A bleed excessively; even small cuts and bruises can be life threatening. Spontaneous bleeding occurs in joints such as elbows, knees, and ankles, producing pain, swelling, and erosion of the bone. Fortunately, bleeding in people with hemophilia A can now be controlled by administering concentrated doses of factor VIII. The inheritance of hemophilia A is illustrated by the family of Queen Victoria of England (Figure 6.8).
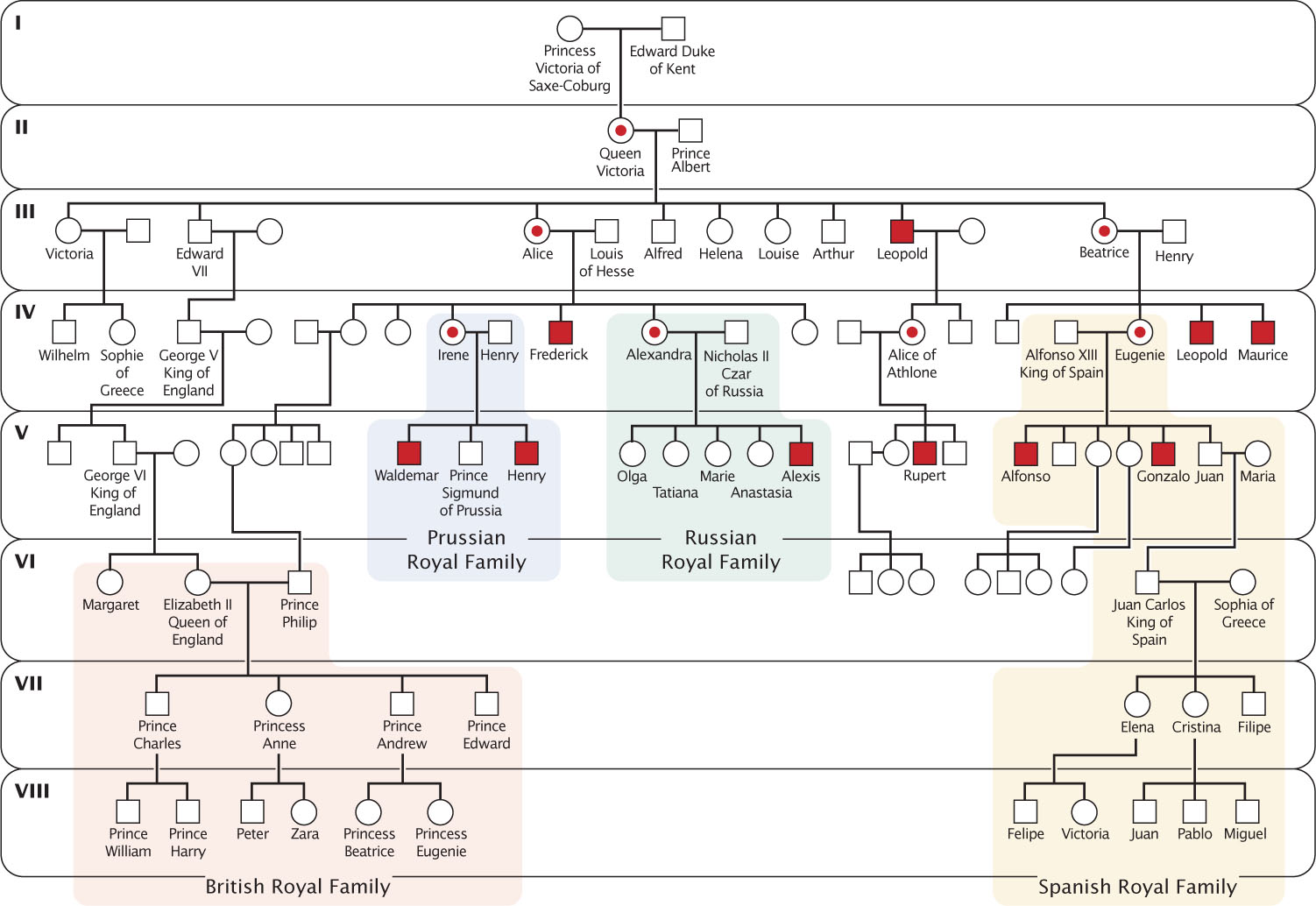
145
X-Linked Dominant Traits
X-linked dominant traits appear in males and females, although they often appear more frequently in females than males. Each person with an X-linked dominant trait must have an affected parent (unless the person possesses a new mutation or the trait has reduced penetrance). X-linked dominant traits do not skip generations (Figure 6.9); affected men pass the trait on to all their daughters and none of their sons, as is seen in the children of I-1 in Figure 6.9. In contrast, affected women (if heterozygous) pass the trait on to about
of their sons and about
of their daughters, as seen in the children of III-6 in the pedigree. As with X-linked recessive traits, a male inherits an X-linked dominant trait only from his mother; the trait is not passed from father to son. This fact is what distinguishes X-linked dominant inheritance from autosomal dominant inheritance, in which a male can inherit the trait from his father. A female, on the other hand, inherits an X chromosome from both her mother and her father; so females can receive an X-linked dominant trait from either parent.
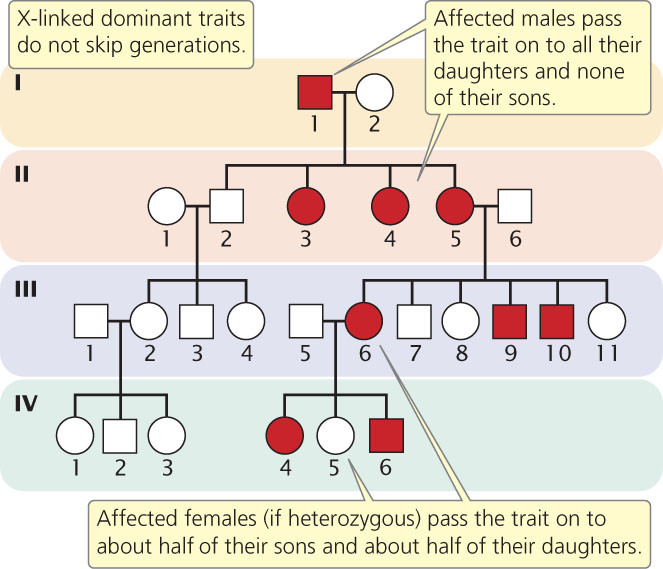
146
CONCEPTS
X-linked dominant traits affect both males and females. Affected males must have affected mothers (unless the males possess a new mutation), and they pass the trait on to all their daughters.
CONCEPT CHECK 4
A male affected with an X-linked dominant trait will have what proportion of offspring affected with the trait?
-
sons and
daughters.
- All sons and no daughtersd
- All daughters and no sons
-
daughters and
sons
An example of an X-linked dominant trait in humans is hypophosphatemia, or familial vitamin-D-resistant rickets. People with this trait have features that superficially resemble those produced by rickets: bone deformities, stiff spines and joints, bowed legs, and mild growth deficiencies. This disorder, however, is resistant to treatment with vitamin D, which normally cures rickets. X-linked hypophosphatemia results from the defective transport of phosphate, especially in cells of the kidneys. People with this disorder excrete large amounts of phosphate in their urine, resulting in low levels of phosphate in the blood and reduced deposition of minerals in the bone. The disorder is treated with high doses of calcitriol (a hormonally active form of vitamin D) and phosphate. As is common with X-linked dominant traits, males with hypophosphatemia are often more severely affected than females.
Y-Linked Traits
Y-linked traits exhibit a specific, easily recognized pattern of inheritance. Only males are affected, and the trait is passed from father to son. If a man is affected, all his male offspring also should be affected, as is the case for II-4, II-6, III-6, and III-10 of the pedigree in Figure 6.10. Y-linked traits do not skip generations. As mentioned in Chapter 4, little genetic information is found on the human Y chromosome. Maleness is one of the few traits in humans that has been shown to be Y-linked. Because each male has only a single Y chromosome, there is only one copy of each Y-linked allele: therefore, Y-linked traits are neither dominant nor recessive.
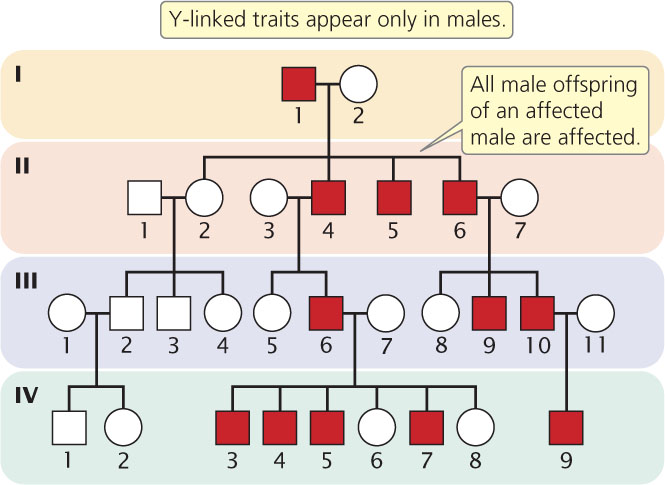
CONCEPTS
Y-linked traits appear only in males and are passed from a father to all his sons.
CONCEPT CHECK 5
What features of a pedigree would distinguish between a Y-linked trait and a trait that is rare, autosomal dominant, and sex-limited to males?
The major characteristics of autosomal recessive, autosomal dominant, X-linked recessive, X-linked dominant, and Y-linked traits are summarized in Table 6.1. ![]() TRY PROBLEM 22
TRY PROBLEM 22
| Autosomal recessive trait |
|
| Autosomal dominant trait |
|
| X-linked recessive trait |
|
| X-linked dominant trait |
|
| Y-linked trait |
|
WORKED PROBLEM
The following pedigree represents the inheritance of a rare disorder in an extended family. What is the most likely mode of inheritance for this disease? (Assume that the trait is fully penetrant.)
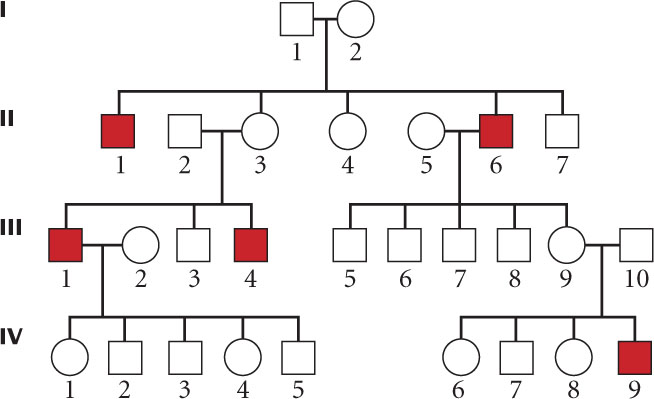
Solution Strategy
What information is required in your answer to the problem?
The most likely mode of inheritance for the trait shown in the pedigree.
What information is provided to solve the problem?
 The pedigree, which includes information about the sex and family relationships of affected individuals.
The pedigree, which includes information about the sex and family relationships of affected individuals.-
The trait is rare.
147
Solution Steps
To answer this question, we should consider each mode of inheritance and determine which, if any, we can eliminate. The trait appears only in males, and so autosomal dominant and autosomal recessive modes of inheritance are unlikely because traits with these modes appear equally in males and females. Additionally, autosomal dominance can be eliminated because some affected persons do not have an affected parent.
The trait is observed only among males in this pedigree, which might suggest Y-linked inheritance. However, for a Y-linked trait, affected men should pass the trait on to all their sons, which is not the case here; II-6 is an affected man who has four unaffected male offspring. We can eliminate Y-linked inheritance.
X-linked dominance can be eliminated because affected men should pass an X-linked dominant trait on to all of their female offspring, and II-6 has an unaffected daughter (III-9).
X-linked recessive traits often appear more commonly in males, and affected males are usually born to unaffected female carriers; the pedigree shows this pattern of inheritance. For an X-linked trait, about half the sons of a heterozygous carrier mother should be affected. II-3 and III-9 are suspected carriers, and about half of their male children (three of five) are affected. Another important characteristic of an X-linked recessive trait is that it is not passed from father to son. We observe no father-to-son transmission in this pedigree. X-linked recessive is therefore the most likely mode of inheritance.
For additional practice, try to determine the mode of inheritance for the pedigrees in Problem 24 at the end of the chapter.