Model Genetic Organism
The Nematode Worm Caenorhabditis elegans
 As we have seen, RNA interference was first demonstrated in the nematode Caenorhabditis elegans when geneticists discovered that they could silence specific genes in this species by injecting the animals with double-
As we have seen, RNA interference was first demonstrated in the nematode Caenorhabditis elegans when geneticists discovered that they could silence specific genes in this species by injecting the animals with double-
282
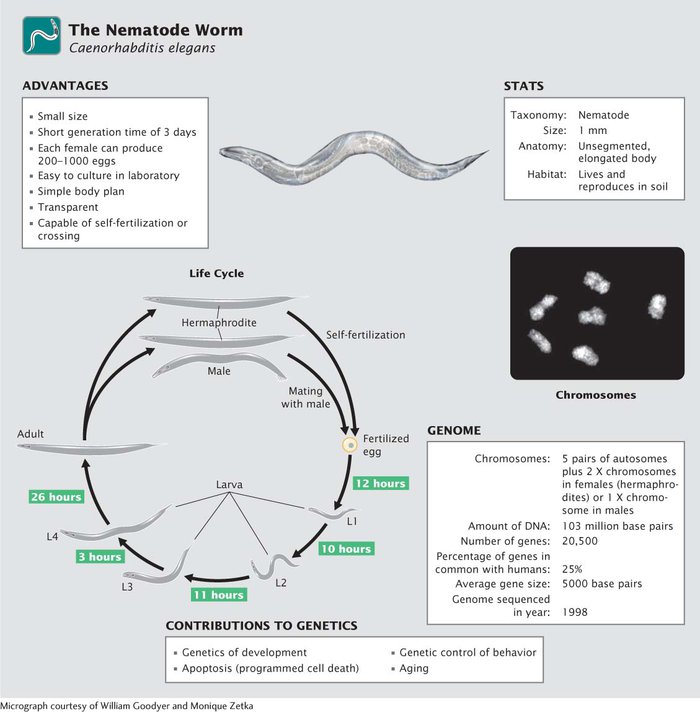
You may be asking what a nematode is and why it is a model genetic organism. Although rarely seen, nematodes are among the most abundant organisms on Earth, inhabiting soils throughout the world. Most of these worms are free living and cause no harm, but a few are important parasites of plants and animals, including humans. Although C. elegans has no economic importance, it has become widely used in genetic studies because of its simple body plan, ease of culture, and high reproductive capacity (Figure 10.26). First introduced to the study of genetics by Sydney Brenner, who formulated plans in 1962 to use C. elegans for the genetic dissection of behavior, this species has made important contributions to the study of development, cell death, aging, and behavior.
Advantages of Caenorhabditis elegans as a model genetic organism
An ideal genetic organism, C. elegans is small, easy to culture, and produces large numbers of offspring. The adult C. elegans is about 1 mm in length. Most investigators grow C. elegans on agar-
Another advantage of C. elegans, particularly for developmental studies, is that the worm is transparent, allowing easy observation of internal development at all stages. It has a simple body structure, with a small, invariant number of somatic cells: 959 cells in a mature hermaphroditic female and 1031 cells in a mature male.
Life cycle
Most mature adults are hermaphrodites, with the ability to produce both eggs and sperm and undergo self-
Eggs are fertilized internally, either by sperm produced by the hermaphrodite or by sperm contributed by a male (see Figure 10.26). The eggs are then laid, and development is completed externally. Approximately 14 hours after fertilization, a larva hatches from the egg and goes through four larval stages—
Genetic techniques
Geneticists began developing plans in 1989 to sequence the genome of C. elegans, and the complete genome sequence was obtained in 1998. Compared with the genomes of most multicellular animals, that of C. elegans, at 103 million base pairs of DNA, is small, which facilitates genomic analysis. The availability of the complete genome sequence provides a great deal of information about gene structure, function, and organization in this species.
Chemical mutagens are routinely used to generate mutations in C. elegans—mutations that are easy to identify and isolate. The ability of hermaphrodites to self-
283
Developmental studies are facilitated by the transparent body of the worm. As mentioned earlier, C. elegans has a small and exact number of somatic cells. Researchers studying the development of C. elegans have meticulously mapped the entire cell lineage of the species, and so the developmental fate of every cell in the adult body can be traced back to the original single-
RNA interference has proved to be an effective tool for turning off genes in C. elegans. Geneticists inject double-
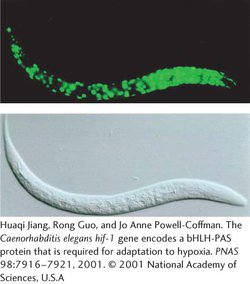
Transgenic worms can be produced by injecting DNA into the ovary, where the DNA becomes incorporated into the oocytes. Geneticists have created a special reporter gene that produces the jellyfish green fluorescent protein (GFP). When this reporter gene is injected into the ovary and becomes inserted into the worm genome, its expression produces GFP, which fluoresces green, allowing the expression of the gene to be easily observed (Figure 10.27).