Coupling and Repulsion
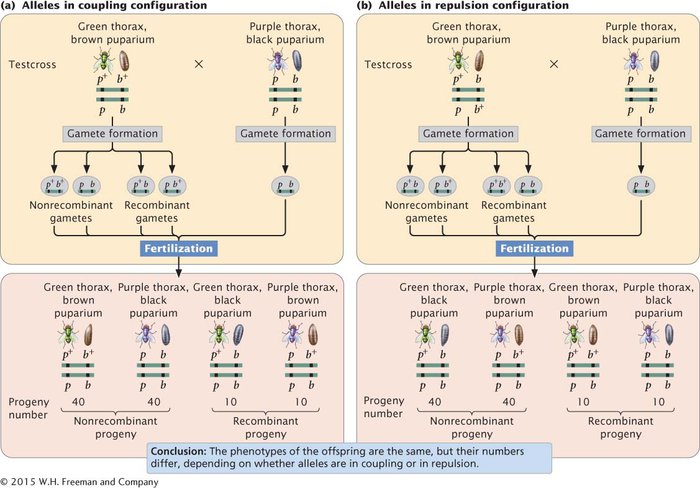
In crosses for linked genes, the arrangement of alleles on the homologous chromosomes is critical in determining the outcome of the cross. For example, consider the inheritance of two genes in the Australian blowfly, Lucilia cuprina. In this species, one locus determines the color of the thorax: a purple thorax (p) is recessive to the normal green thorax (p+). A second locus determines the color of the puparium: a black puparium (b) is recessive to the normal brown puparium (b+). The loci for thorax color and puparium color are located close together on the same chromosome. Suppose that we test-
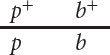
This arrangement, in which wild-
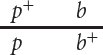
This arrangement, in which each chromosome contains one wild-
When the alleles are in the coupling configuration, the most numerous progeny types are those with a green thorax and brown puparium and those with a purple thorax and black puparium (Figure 5.7a). However, when the alleles of the heterozygous parent are in repulsion, the most numerous progeny types are those with a green thorax and black puparium and those with a purple thorax and brown puparium (Figure 5.7b). Notice that the genotypes of the parents in Figure 5.7a and 5.7b are the same (p+pb+b × pp bb) and that the dramatic difference in the phenotypic ratios of the progeny in the two crosses results entirely from the configuration—
123
CONCEPTS
In a cross, the arrangement of linked alleles on the chromosomes is critical for determining the outcome. When two wild-
 CONCEPT CHECK 2
CONCEPT CHECK 2
The following testcross produces the progeny shown: Aa Bb × aa bb → 10 Aa Bb, 40 Aa bb, 40 aa Bb, 10 aa bb. Were the A and B alleles in the Aa Bb parent in coupling or in repulsion?
Repulsion
CONNECTING CONCEPTS
Relating Independent Assortment, Linkage, and Crossing Over
We have now considered three possible situations among genes at different loci. First, the genes may be located on different chromosomes; in this case, they exhibit independent assortment and combine randomly when gametes are formed. An individual heterozygous at two loci (Aa Bb) produces four types of gametes (A B, a b, A b, and a B) in equal proportions: two types of nonrecombinants and two types of recombinants. In a testcross, these gametes will result in four types of progeny in equal proportions (Table 5.1).
Second, the genes may be completely linked—

produces only nonrecombinant gametes containing alleles A B or a b. The alleles do not assort into new combinations such as A b or a B. In a testcross, completely linked genes will produce only two types of progeny, both nonrecombinants, in equal proportions (see Table 5.1).
The third situation, incomplete linkage, is intermediate between the two extremes of independent assortment and complete linkage. Here, the genes are physically linked on the same chromosome, which prevents independent assortment. However, occasional crossovers break up the linkage and allow the genes to recombine. With incomplete linkage, an individual heterozygous at two loci produces four types of gametes—
| Situation | Progeny of Testcross | |
|---|---|---|
| Independent assortment | Aa Bb (nonrecombinant) | 25% |
| aa bb (nonrecombinant) | 25% | |
| Aa bb (recombinant) | 25% | |
| aa Bb (recombinant) | 25% | |
| Complete linkage (genes in coupling) | Aa Bb (nonrecombinant) | 50% |
| aa bb (nonrecombinant) | 50% | |
| Linkage with some crossing over (genes in coupling) |  |
more than 50% |
 |
less than 50% |
124
Earlier in the chapter, the term recombination was defined as the sorting of alleles into new combinations. We’ve now considered two types of recombination that differ in their mechanisms. Interchromosomal recombination takes place between genes located on different chromosomes. It arises from independent assortment—