Concept 17.2: Speciation Is a Natural Consequence of Population Subdivision
Not all evolutionary changes result in new species. A single lineage may change over time without giving rise to a new species. Speciation requires the interruption of gene flow within a species whose members formerly exchanged genes. But if a genetic change prevents reproduction between individuals of a species, how can such a change spread through a species in the first place?
Incompatibilities between genes can produce reproductive isolation
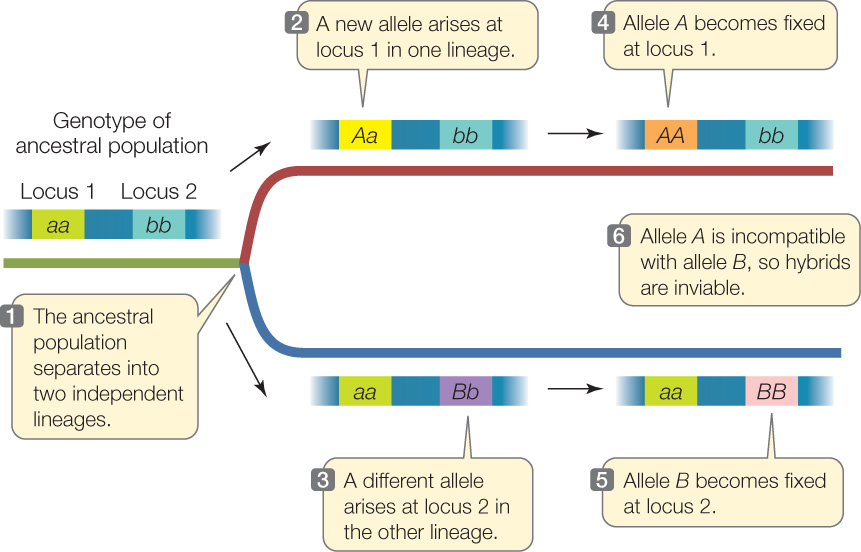
If a new allele that causes reproductive incompatibility arises in a population, it cannot spread through the population because no other individuals will be reproductively compatible with the individual that carries the new allele. So how can one reproductively cohesive lineage ever split into two reproductively isolated species? Several early geneticists, including Theodosius Dobzhansky and Hermann Joseph Muller, developed a genetic model to explain this apparent conundrum (FIGURE 17.3).
The Dobzhansky–Muller model is quite simple. First, assume that a single ancestral population is subdivided into two daughter populations (by the formation of a new mountain range, for instance), which then evolve as independent lineages. In one of the descendant lineages, a new allele (A) arises and becomes fixed (see Figure 17.3). In the other population, another new allele (B) becomes fixed at a different gene locus. Neither new allele at either locus results in any loss of reproductive compatibility. However, the two new forms of these two different genes have never occurred together in the same individual or population. Recall that the products of many genes must work together in an organism. It is possible that the new protein forms encoded by the two new alleles will not be compatible with each other. If individuals from the two lineages come back together after these genetic changes, they may still be able to interbreed. However, the hybrid offspring may have a new combination of genes that is functionally inferior, or even lethal. This will not happen with all new combinations of genes, but over time, isolated lineages will accumulate many allele differences at many gene loci. Some combinations of these differentiated genes will not function well together in hybrids. Thus genetic incompatibility between the two isolated populations will develop over time.

Go to ANIMATED TUTORIAL 17.1 Speciation Simulation
PoL2e.com/at17.1
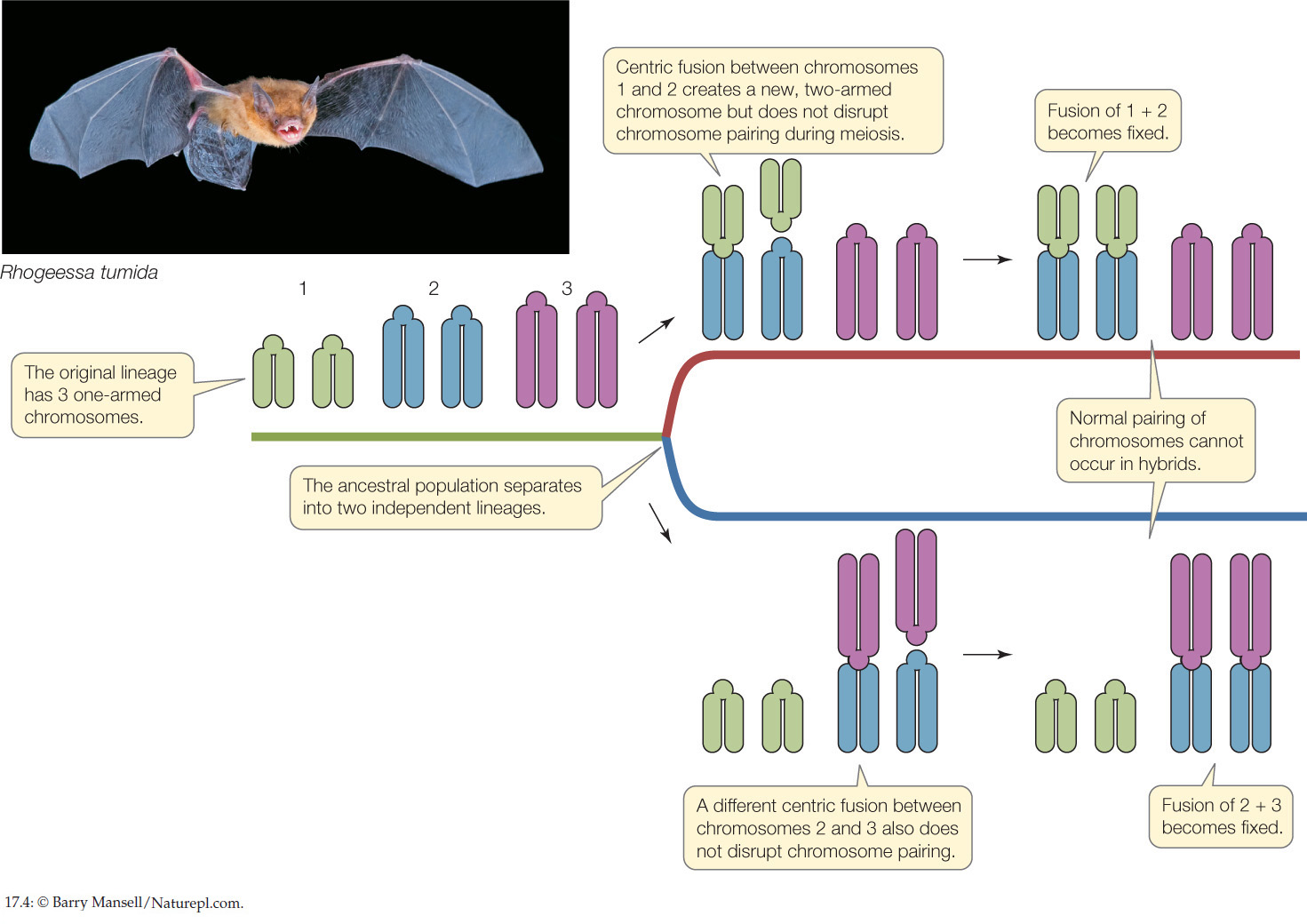
Many empirical and experimental examples support the Dobzhansky–Muller model. This model works not only for pairs of individual genes but also for some kinds of chromosomal rearrangements. Bats of the genus Rhogeessa, for example, exhibit considerable variation in centric fusions of their chromosomes. The chromosomes of the various species contain the same basic chromosomal arms, but in some species two acrocentric (one-armed) chromosomes have fused at the centromere to form larger, metacentric (two-armed) chromosomes. A polymorphism in centric fusion causes few, if any, problems in meiosis because the respective chromosomes can still align and assort normally. Therefore a given centric fusion can become fixed in a lineage. However, if a different centric fusion becomes fixed in a second lineage, then hybrids between individuals of each lineage will not be able to produce normal gametes in meiosis (FIGURE 17.4). Most of the closely related species of Rhogeessa display different combinations of these centric fusions and are thereby reproductively isolated from one another.
Reproductive isolation develops with increasing genetic divergence
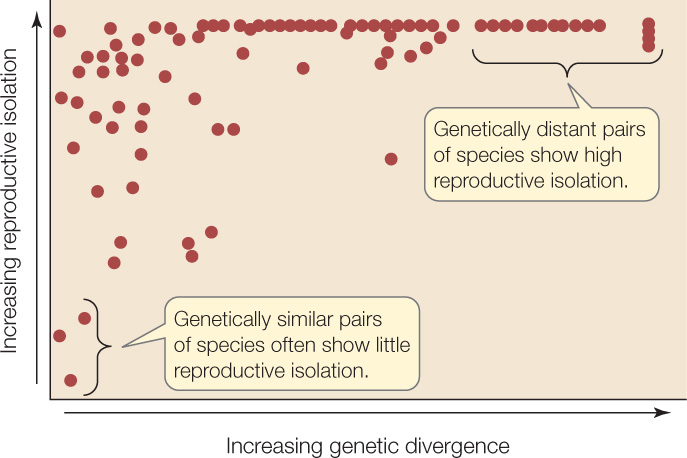
As pairs of species diverge genetically, they become increasingly reproductively isolated (FIGURE 17.5). Both the rate at which reproductive isolation develops and the mechanisms that produce it vary from group to group. Reproductive incompatibility has been shown to develop gradually in many groups of plants, animals, and fungi, reflecting the slow pace at which incompatible genes accumulate in each lineage. In some cases, complete reproductive isolation may take millions of years. In other cases (as with the chromosomal fusions of Rhogeessa described above), reproductive isolation can develop over just a few generations.
Partial reproductive isolation has evolved in strains of Phlox drummondii artificially isolated by humans. In 1835, Thomas Drummond, after whom this species of garden plant is named, collected seeds in Texas and distributed them to nurseries in Europe. The European nurseries established more than 200 true-breeding strains of P. drummondii that differed in flower size, flower color, and plant growth form. The breeders did not select directly for reproductive incompatibility between strains, but in subsequent experiments in which strains were crossed and seed production was measured and compared, biologists found that reproductive compatibility between strains had been reduced by 14 to 50 percent, depending on the cross—even though the strains had been isolated from one another for less than two centuries.
347
CHECKpoint CONCEPT 17.2
- The Dobzhansky–Muller model suggests that divergence among alleles at different gene loci leads to genetic incompatibility between species. Why is genetic incompatibility between two alleles at the same locus considered less likely?
- Why do some combinations of chromosomal centric fusions cause problems in meiosis? Can you diagram what would happen at meiosis in a hybrid of the divergent lineages shown in Figure 17.4?
- Assume that the reproductive isolation seen in Phlox strains results from lethal combinations of incompatible alleles at several loci among the various strains. Given this assumption, why might the reproductive isolation seen among these strains be partial rather than complete?
We have now seen how splitting an ancestral population leads to genetic divergence in the two descendant lineages. Next we will consider ways in which the descendant lineages could have become separated in the first place.
348