Concept 20.2: Major Lineages of Eukaryotes Diversified in the Precambrian
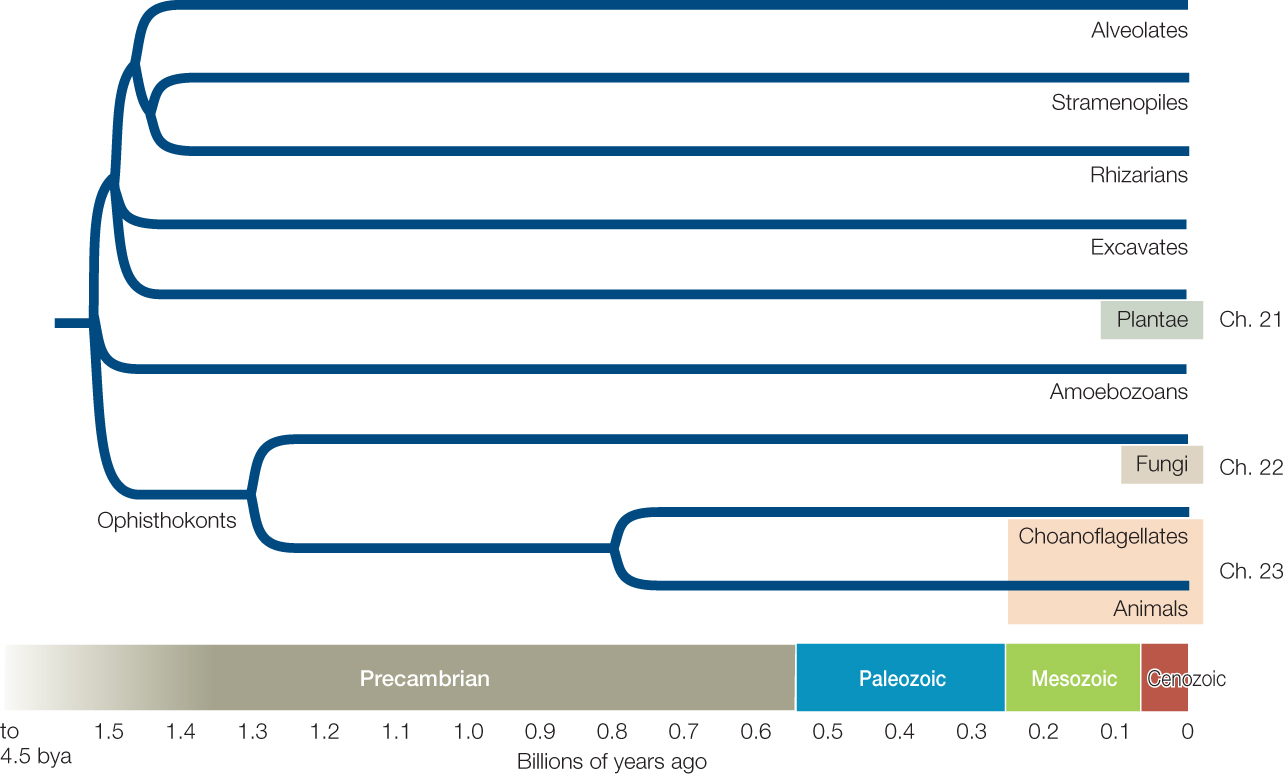
Most eukaryotes can be classified in one of eight major clades that began to diversify about 1.5 billion years ago: alveolates, excavates, stramenopiles, plants, rhizarians, amoebozoans, fungi, and animals (FIGURE 20.3). Plants, fungi, and animals each have close protist relatives (such as the choanoflagellate relatives of animals), which we will discuss along with those major multicellular eukaryote groups in Chapters 21–23.
Each of the five major groups of protist eukaryotes covered in this chapter consists of organisms with enormously diverse body forms and nutritional lifestyles. Some protists are motile, whereas others do not move. Some protists are photosynthetic, whereas others are heterotrophic. Most protists are unicellular, but some are multicellular. Most protists are microscopic, but a few are huge (giant kelps, for example, can grow to half the length of a football field). We refer to the unicellular species of protists as microbial eukaryotes, but keep in mind that there are large, multicellular protists as well.
Multicellularity has arisen dozens of times across the evolutionary history of eukaryotes. Four of the origins of multicellularity resulted in large organisms that are familiar to most people: plants, animals, fungi, and brown algae (the last are a group of stramenopiles). In addition, there are dozens of smaller and less familiar groups among the eukaryotes that include multicellular species. Recent experimental studies have shown that artificial selection for multicellularity can produce repeated, convergent evolution of multicellular forms over just a few months in some normally unicellular eukaryotic species. In addition, many unicellular species retain individual identities but nonetheless associate in large multicellular colonies. There is a near-continuum between fully integrated, multicellular organisms on the one hand and loosely integrated multicellular colonies of cells on the other. Biologists do not always agree on where to draw the line between the two.
406
Biologists used to classify protists largely on the basis of their life histories and reproductive features (see Concept 20.3). In recent years, however, electron microscopy and gene sequencing have revealed many new patterns of evolutionary relatedness among these groups. Analyses of slowly evolving gene sequences are making it possible to explore evolutionary relationships among eukaryotes in ever greater detail and with greater confidence. Nonetheless, some substantial areas of uncertainty remain, and lateral gene transfer may complicate efforts to reconstruct the evolutionary history of protists (as was also true for prokaryotes; see Concept 19.1). Today we recognize great diversity among the many distantly related protist clades.
Alveolates have sacs under their cell membranes
Alveolates are so named because they possess sacs, called alveoli, just beneath their cell membranes, which may play a role in supporting the cell surface. All alveolates are unicellular, and most are photosynthetic, but they are diverse in body form. The groups considered in detail here are the dinoflagellates, apicomplexans, and ciliates.
Dinoflagellates
Most dinoflagellates are marine and photosynthetic; they are important primary producers of organic matter in the oceans. Although fewer species of dinoflagellates live in fresh water, individuals can be abundant in freshwater environments. The dinoflagellates are of great ecological, evolutionary, and morphological interest. A distinctive mixture of photosynthetic and accessory pigments gives their chloroplasts a golden brown color. Some dinoflagellate species cause red tides, as discussed at the start of this chapter. Other species are photosynthetic endosymbionts that live within the cells of other organisms, including invertebrate animals (such as corals; see Figure 20.19) and other marine protists (see Figure 20.11A). Some are nonphotosynthetic and live as parasites within various marine organisms.
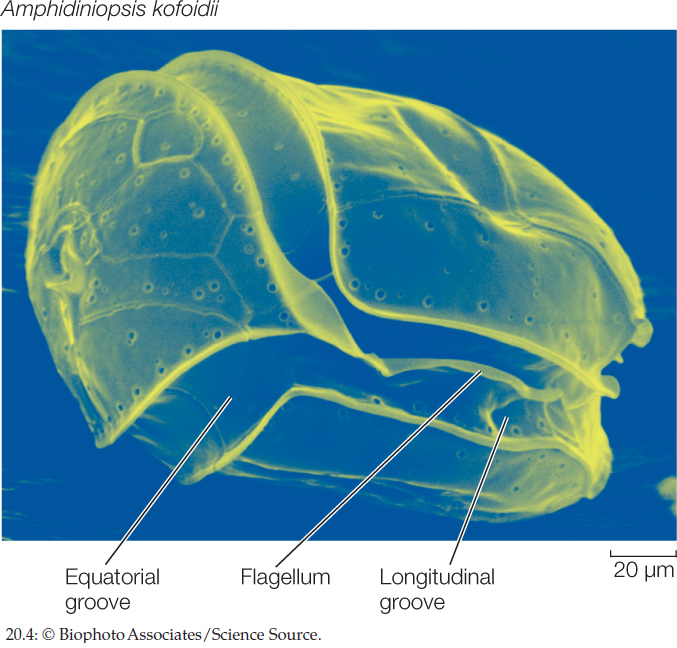
Dinoflagellates have a distinctive appearance. They generally have two flagella, one in an equatorial groove around the cell, the other starting near the same point as the first and passing down a longitudinal groove before extending into the surrounding medium (FIGURE 20.4). Some dinoflagellates can take on different forms, including amoeboid ones, depending on environmental conditions. It has been claimed that the dinoflagellate Pfiesteria piscicida can occur in at least two dozen distinct forms, although this claim is highly controversial. In any case, this remarkable dinoflagellate is harmful to fish and can, when present in great numbers, both stun and feed on them.

Go to MEDIA CLIP 20.1 A Dinoflagellate Shows Off Its Flagellum
PoL2e.com/mc20.1
Apicomplexans
The exclusively parasitic apicomplexans derive their name from the apical complex, a mass of organelles contained in the apical end (the tip) of the cell. These organelles help the apicomplexan invade its host’s tissues. For example, the apical complex enables Plasmodium, the causative agent of malaria, to enter its target cells in the human body after transmission by a mosquito.
Like many obligate parasites, apicomplexans have elaborate life cycles featuring asexual and sexual reproduction by a series of very dissimilar life stages. In many species, these life stages are associated with two different types of host organisms, as is the case with Plasmodium. Another apicomplexan, Toxoplasma, alternates between cats and rats to complete its life cycle. A rat infected with Toxoplasma loses its fear of cats, which makes it more likely to be eaten by, and thus transfer the parasite to, a cat.
Ciliates
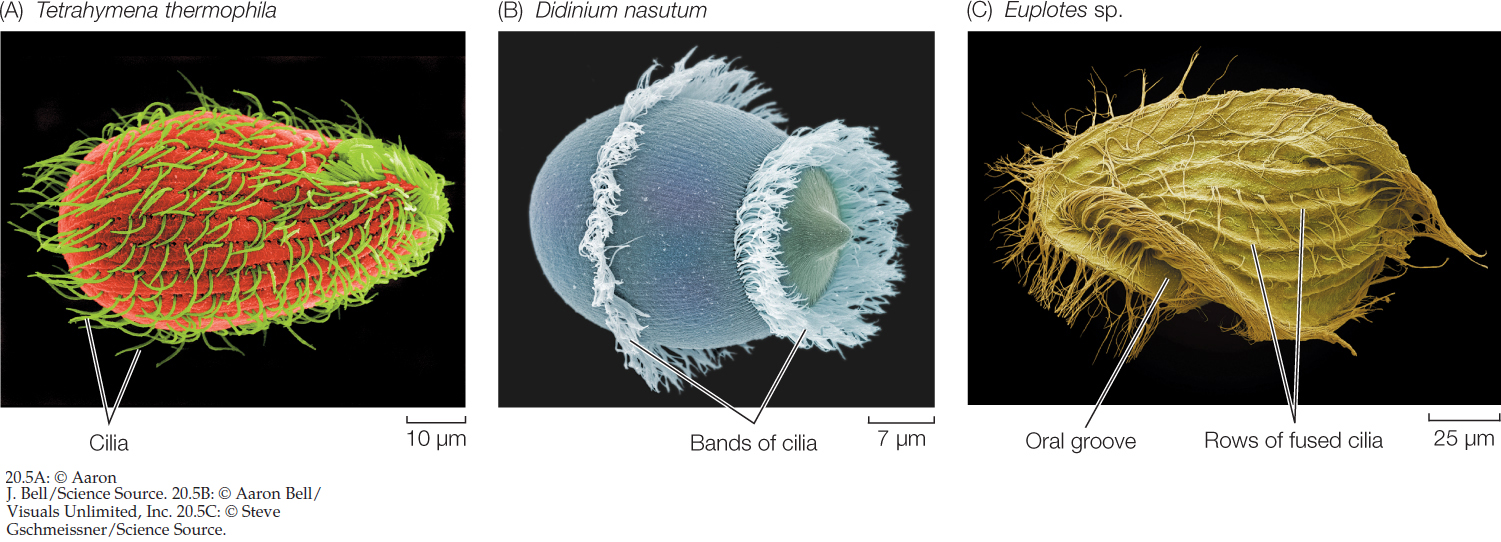
The ciliates are named for their numerous hairlike cilia, which are shorter than, but otherwise identical to, eukaryotic flagella. The ciliates are much more complex in body form than are most other unicellular eukaryotes (FIGURE 20.5). Their definitive characteristic is the possession of two types of nuclei (whose roles we will describe in Concept 20.3 when we discuss protist reproduction). Almost all ciliates are heterotrophic, although a few contain photosynthetic endosymbionts.
407
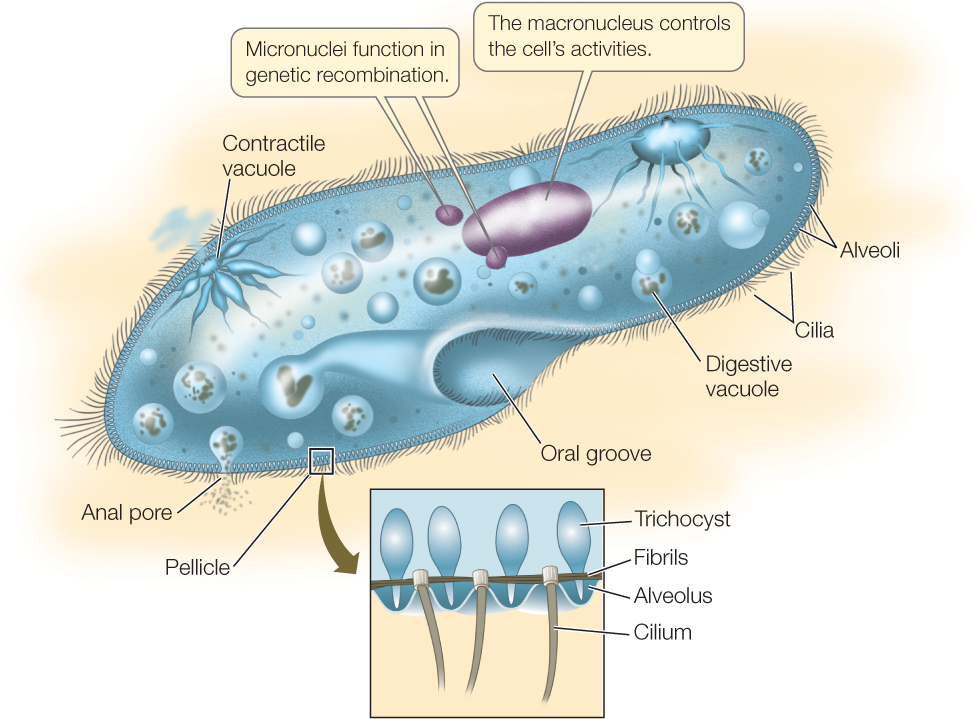
Paramecium, a frequently studied ciliate genus, exemplifies the complex structure and behavior of ciliates (FIGURE 20.6). The slipper-shaped cell is covered by an elaborate pellicle, a structure composed principally of an outer membrane and an inner layer of closely packed, membrane-enclosed sacs (the alveoli) that surround the bases of the cilia. Defensive organelles called trichocysts are also present in the pellicle. In response to a threat, a microscopic explosion expels the trichocysts in a few milliseconds, and they emerge as sharp darts, driven forward at the tip of a long, expanding filament.
Go to ACTIVITY 20.1 Anatomy of a Paramecium
PoL2e.com/ac20.1
The cilia provide Paramecium with a form of locomotion that is generally more precise than locomotion by flagella or pseudopods. A Paramecium can coordinate the beating of its cilia to propel itself either forward or backward in a spiraling manner. It can also back off swiftly when it encounters a barrier or a negative stimulus. The coordination of ciliary beating is probably the result of a differential distribution of ion channels in the cell membrane near the two ends of the cell.
Organisms living in fresh water are hypertonic to their environment. Many freshwater protists, including Paramecium, address this problem by means of specialized contractile vacuoles that excrete the excess water the organisms constantly take in by osmosis. The excess water collects in the contractile vacuoles, which then contract and expel the water from the cell.
Paramecium and many other protists engulf solid food by endocytosis, forming a digestive vacuole within which the food is digested. Smaller vesicles containing digested food pinch away from the digestive vacuole and enter the cytoplasm. These tiny vesicles provide a large surface area across which the products of digestion can be absorbed by the rest of the cell.

Go to ANIMATED TUTORIAL 20.2 Digestive Vacuoles
PoL2e.com/at20.2
LINK
The role of a pathogen’s cell surface recognition molecules in the mammalian immune response is covered in Chapter 39
408
Stramenopiles typically have two unequal flagella, one with hairs
A morphological synapomorphy of most stramenopiles is the possession of rows of tubular hairs on the longer of their two flagella. Some stramenopiles lack flagella, but they are descended from ancestors that possessed flagella. The stramenopiles include the diatoms and the brown algae, which are photosynthetic, and the oomycetes, which are not.
Diatoms
All of the diatoms are unicellular, although some species associate in filaments. Many have sufficient carotenoids in their chloroplasts to give them a yellow or brownish color. All of them synthesize carbohydrates and oils as photosynthetic storage products. Diatoms lack flagella except in male gametes.
Architectural magnificence on a microscopic scale is the hallmark of the diatoms. Almost all diatoms deposit silica (hydrated silicon dioxide) in their cell walls. The cell wall of a diatom is constructed in two pieces, with the top overlapping the bottom like the top of a petri dish. The silica-impregnated walls have intricate patterns unique to each species (FIGURE 20.7). Despite their remarkable morphological diversity, all diatoms are symmetrical—either bilaterally (with “right” and “left” halves) or radially (with the type of symmetry possessed by a circle).


Go to MEDIA CLIP 20.2 Diatoms in Action
PoL2e.com/mc20.2
Diatoms reproduce both sexually and asexually. Asexual reproduction by binary fission is somewhat constrained by the stiff cell wall. Both the top and bottom of the “petri dish” become tops of new “dishes” without changing appreciably in size. As a result, the new cell made from the former bottom is smaller than the parent cell. If this process continued indefinitely, one cell line would simply vanish, but sexual reproduction largely solves this potential problem. Gametes are formed, shed their cell walls, and fuse. The resulting zygote then grows substantially in size before a new cell wall is laid down.
Diatoms are found in all the oceans and are frequently present in great numbers. They are major photosynthetic producers in coastal waters and are among the dominant organisms in the dense “blooms” of phytoplankton that occasionally appear in the open ocean (see Concept 20.4). Diatoms are also common in fresh water and even occur on the wet surfaces of terrestrial mosses.
Brown Algae
The brown algae obtain their namesake color from the carotenoid fucoxanthin, which is abundant in their chloroplasts. The combination of this yellow-orange pigment with the green of chlorophylls a and c yields a brownish tinge. All brown algae are multicellular, and some are extremely large. Giant kelps, such as those of the genus Macrocystis, may be up to 60 meters long.

Go to MEDIA CLIP 20.3 A Kelp Forest
PoL2e.com/mc20.3
The brown algae are almost exclusively marine. They are composed either of branched filaments (FIGURE 20.8A) or of leaflike growths (FIGURE 20.8B). Some float in the open ocean. The most famous example is the genus Sargassum, which forms dense mats in the Sargasso Sea in the mid-Atlantic. Most brown algae, however, attach themselves to rocks near the shore. A few thrive only where they are regularly exposed to heavy surf. All of the attached forms develop a specialized structure, called a holdfast, that literally glues them to the rocks. The “glue” of the holdfast is alginic acid, a gummy polymer of sugar acids found in the walls of many brown algal cells. In addition to its function in holdfasts, it cements algal cells and filaments together. It is harvested and used by humans as an emulsifier in ice cream, cosmetics, and other products.

Oomycetes
The oomycetes consist in large part of the water molds and their terrestrial relatives, such as the downy mildews. Water molds are filamentous and stationary. They are absorptive heterotrophs—that is, they secrete enzymes that digest large food molecules into smaller molecules that they can absorb. They are all aquatic and saprobic—meaning they feed on dead organic matter. If you have seen a whitish, cottony mold growing on dead fish or dead insects in water, it was probably a water mold of the common genus Saprolegnia (FIGURE 20.9).

409
Some other oomycetes, such as the downy mildews, are terrestrial. Although most of the terrestrial oomycetes are harmless or helpful decomposers of dead matter, a few are plant parasites that attack crops such as avocados, grapes, and potatoes.
Oomycetes were once classified as fungi. However, we now know that their similarity to fungi is only superficial, and that the oomycetes are more distantly related to the fungi than are many other eukaryote groups, including humans (see Figure 20.3). For example, the cell walls of oomycetes are typically made of cellulose, whereas those of fungi are made of chitin.
Rhizarians typically have long, thin pseudopods
The three primary groups of rhizarians—cercozoans, foraminiferans, and radiolarians—are unicellular and mostly aquatic. These organisms typically have long, thin pseudopods that contrast with the broader, lobelike pseudopods of the more familiar amoebozoans. The rhizarians have contributed to ocean sediments, some of which have become terrestrial features in the course of geological history.
Cercozoans
The cercozoans are a diverse group with many forms and habitats. Some are aquatic, and others live in soil. One group of cercozoans possesses chloroplasts derived from a green alga by secondary endosymbiosis, and those chloroplasts contain a trace of the alga’s nucleus.
Foraminiferans
Some foraminiferans secrete external shells of calcium carbonate (FIGURE 20.10). These shells have accumulated over time to produce much of the world’s limestone. Foraminiferans live as plankton (drifing with the curent) or on the seafloor. Living foraminiferans have been found 10,896 meters down in the western Pacific’s Challenger Deep—the deepest point in the world’s oceans. At that depth, however, they cannot secrete normal shells because the surrounding water is too poor in calcium carbonate.

In living planktonic foraminiferans, long, threadlike, branched pseudopods extend through numerous microscopic openings in the shell and interconnect to create a sticky network, which the foraminiferans use to catch smaller plankton. In some foraminiferan species, the pseudopods provide locomotion.
410
Radiolarians
Radiolarians are recognizable by their thin, stiff pseudopods, which are reinforced by microtubules (FIGURE 20.11A). These pseudopods greatly increase the surface area of the cell, and they help the cell stay afloat in its marine environment.

Radiolarians also are immediately recognizable by their distinctive radial symmetry. Almost all radiolarian species secrete glassy endoskeletons (internal skeletons). The skeletons of the different species are as varied as snowflakes, and many have elaborate geometric designs (FIGURE 20.11B). A few radiolarians are among the largest of the unicellular eukaryotes, measuring several millimeters in diameter.
Excavates began to diversify about 1.5 billion years ago
The excavates include a number of diverse groups that began to split from one another soon after the origin of eukaryotes. Several groups of excavates lack mitochondria, an absence that once led to the view that these groups might represent early-diverging eukaryotes that diversified before the evolution of mitochondria. However, the discovery of nuclear genes normally associated with mitochondria in these organisms suggests that the absence of mitochondria is a derived condition. In other words, ancestors of these excavate groups probably possessed mitochondria that were lost or reduced over the course of evolution. The existence of these organisms today shows that eukaryotic life is possible without mitochondria.
Diplomonads and Parabasalids
The diplomonads and parabasalids are unicellular and lack mitochondria. Giardia lamblia, a diplomonad, is a familiar parasite that contaminates water supplies and causes the intestinal disease giardiasis. This tiny organism contains two nuclei bounded by nuclear envelopes, and it has a cytoskeleton and multiple flagella (FIGURE 20.12A).
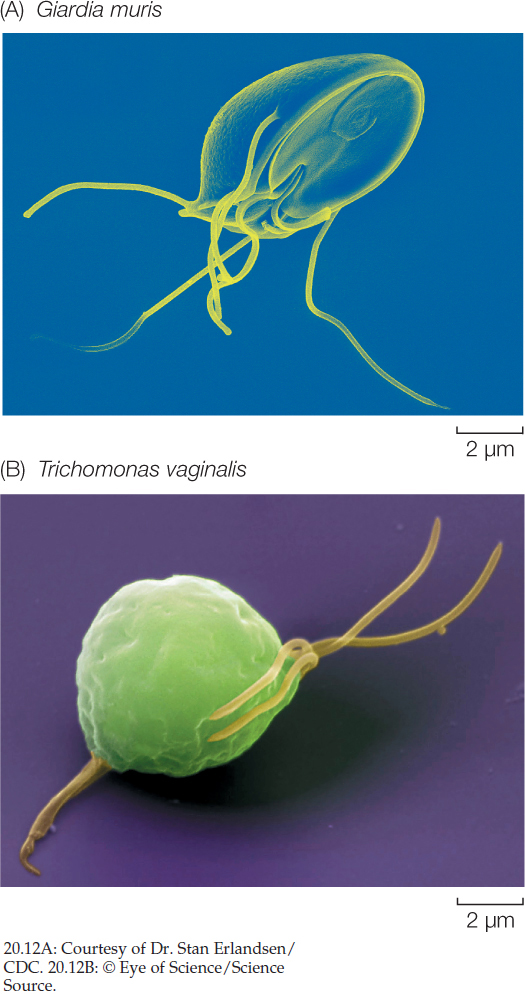
In addition to flagella and a cytoskeleton, the parabasalids have undulating membranes that also contribute to the cell’s locomotion. Trichomonas vaginalis (FIGURE 20.12B) is a parabasalid responsible for a sexually transmitted disease (trichomoniasis) in humans. Infection of the male urethra, where it may occur without symptoms, is less common than infection of the vagina.
Heteroloboseans
The constantly changing amoeboid body form appears in several protist groups that are only distantly related to one another. The body forms of heteroloboseans, for example, resemble those of loboseans, an amoebozoan group that is not at all closely related to heteroloboseans. Amoebas of the free-living heterolobosean genus Naegleria usually have a two-stage life cycle in which one stage has amoeboid cells and the other flagellated cells. Some amoebas of this genus can enter the human body and cause a fatal disease of the nervous system.
Euglenids and Kinetoplastids
The euglenids and kinetoplastids together constitute a clade of unicellular excavates with flagella. Their mitochondria contain distinctive, disc-shaped cristae, and their flagella contain a crystalline rod not found in other organisms. They reproduce primarily asexually by binary fission.
The flagella of euglenids arise from a pocket at the anterior end of the cell. Spiraling strips of proteins under the cell membrane control the cell’s shape. Some euglenids are photosynthetic.
411
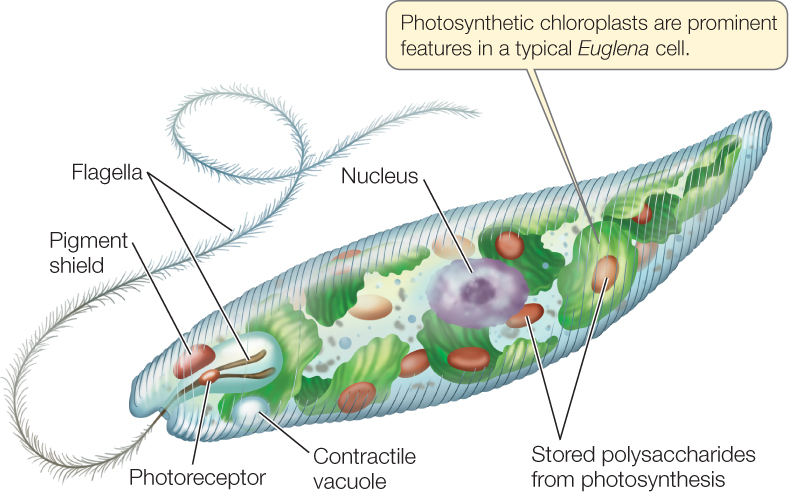
FIGURE 20.13 depicts a cell of the genus Euglena. Like most other euglenids, this common freshwater organism has a complex cell structure. It propels itself through the water with the longer of its two flagella, which may also serve as an anchor to hold the organism in place. The second flagellum is often rudimentary.

Go to MEDIA CLIP 20.4 Euglenids
PoL2e.com/mc20.4
The euglenids have diverse nutritional requirements. Many species are always heterotrophic. Other species are fully autotrophic in sunlight, using chloroplasts to synthesize organic compounds through photosynthesis. When kept in the dark, these euglenids lose their photosynthetic pigment and begin to feed exclusively on dissolved organic material in the water around them. Such a “bleached” Euglena resynthesizes its photosynthetic pigment when it is returned to the light and becomes autotrophic again. But Euglena cells treated with certain antibiotics or mutagens lose their photosynthetic pigment completely; neither they nor their descendants are ever autotrophs again. However, those descendants function well as heterotrophs.
The kinetoplastids are unicellular parasites with two flagella and a single, large mitochondrion. That mitochondrion contains a kinetoplast, a unique structure housing multiple circular DNA molecules and associated proteins. Some of these DNA molecules encode “guide proteins” that edit mRNA within the mitochondrion.
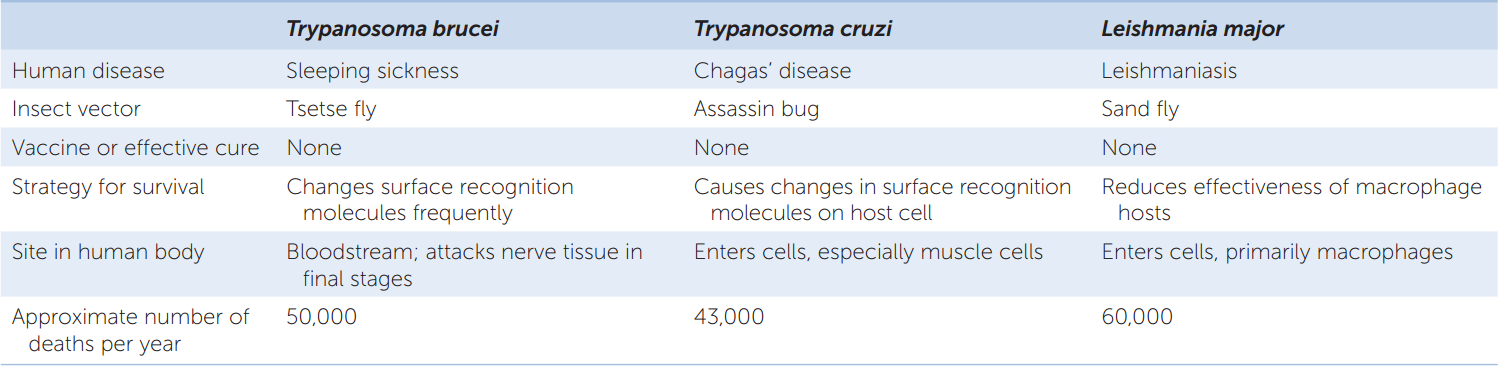
The kinetoplastids include several medically important species of pathogenic trypanosomes (TABLE 20.1). Some of these organisms are able to change their cell surface recognition molecules frequently, allowing them to evade our best attempts to kill them and thus eradicate the diseases they cause.
412
Amoebozoans use lobe-shaped pseudopods for locomotion
Amoebozoans appear to have diverged from other eukaryotes about 1.5 billion years ago (see Figure 20.3). It is not yet clear if they are more closely related to opisthokonts (including fungi and animals) or to other major groups of eukaryotes.
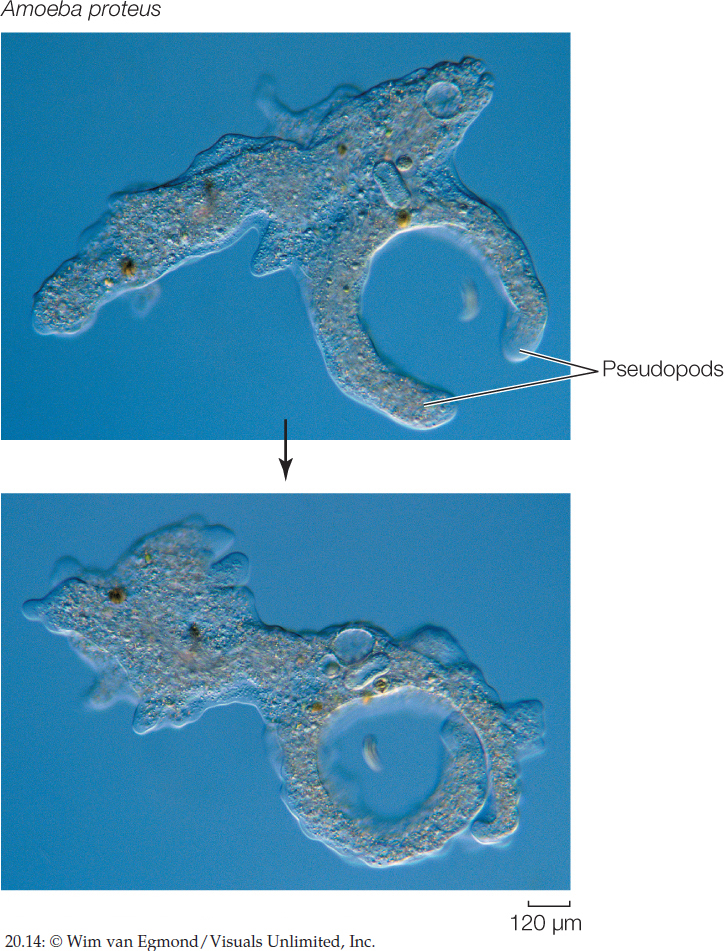
The lobe-shaped pseudopods of amoebozoans (FIGURE 20.14) are a hallmark of the amoeboid body form. Amoebozoan pseudopods differ in form and function from the slender pseudopods of rhizarians. We consider three amoebozoan groups here: the loboseans and two groups known as slime molds.

Go to MEDIA CLIP 20.5 Amoeboid Movement
PoL2e.com/mc20.5
Loboseans
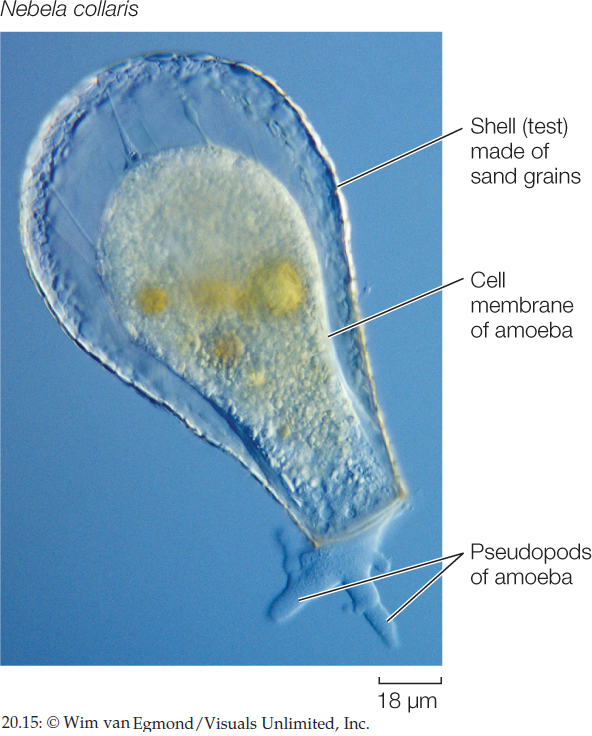
Loboseans are small amoebozoans that feed on other small organisms and particles of organic matter by phagocytosis, engulfing them with pseudopods. Many loboseans are adapted for life on the bottoms of lakes, ponds, and other bodies of water. Their creeping locomotion and their manner of engulfing food particles fit them for life close to a relatively rich supply of sedentary organisms or organic particles. Most loboseans exist as predators, parasites, or scavengers. Members of one group of loboseans, the testate amoebas, live inside shells. Some of these amoebas produce casings by gluing sand grains together (FIGURE 20.15). Other testate amoebas have shells secreted by the organism itself.
Plasmodial Slime Molds
If the nucleus of an amoeba began rapid mitotic division, accompanied by a tremendous increase in cytoplasm and organelles but no cytokinesis (division of the cytoplasm), the resulting organism would resemble the multinucleate mass of a plasmodial slime mold. During its vegetative (feeding, nonreproductive) stage, a plasmodial slime mold is a wall-less mass of cytoplasm with numerous diploid nuclei. This mass streams very slowly over its substrate in a remarkable network of strands called a plasmodium (FIGURE 20.16A). The plasmodium of such a slime mold is an example of a coenocyte: many nuclei enclosed in a single cell membrane. The outer cytoplasm of the plasmodium (closest to the environment) is normally less fluid than the interior cytoplasm and thus provides some structural rigidity.


Go to MEDIA CLIP 20.6 Plasmodial Slime Mold Growth
PoL2e.com/mc20.6
Plasmodial slime molds provide a dramatic example of movement by cytoplasmic streaming. The outer cytoplasmic region of the plasmodium becomes more fluid in places, and cytoplasm rushes into those areas, stretching the plasmodium. This streaming somehow reverses its direction every few minutes as cytoplasm rushes into a new area and drains away from an older one, moving the plasmodium over its substrate. Sometimes an entire wave of plasmodium moves across a surface, leaving strands behind. Microfilaments and a contractile protein interact to produce the streaming movement. As it moves, the plasmodium engulfs food particles by endocytosis—predominantly bacteria, yeasts, spores of fungi, and other small organisms as well as decaying animal and plant remains.
413
A plasmodial slime mold can grow almost indefinitely in its plasmodial stage as long as the food supply is adequate and other conditions, such as moisture and pH, are favorable. If conditions become unfavorable, however, one of two things can happen. First, the plasmodium can form an irregular mass of hardened cell-like components. This resting structure rapidly becomes a plasmodium again when favorable conditions are restored.
Alternatively, the plasmodium can transform itself into spore-bearing fruiting structures (FIGURE 20.16B). These stalked or branched structures rise from heaped masses of plasmodium. They derive their rigidity from walls that form and thicken between their nuclei. The diploid nuclei of the plasmodium divide by meiosis as the fruiting structure develops. One or more knobs, called sporangia, develop on the end of the stalk. Within a sporangium, haploid nuclei become surrounded by walls to form spores. Eventually, as the fruiting structure dries, it sheds its spores.
The spores germinate into wall-less, haploid cells called swarm cells, which can either divide mitotically to produce more haploid swarm cells or function as gametes. Swarm cells can live as separate individual cells that move by means of flagella or pseudopods, or they can become walled and resistant resting cysts when conditions are unfavorable. When conditions improve again, the cysts release swarm cells. Two swarm cells can also fuse to form a diploid zygote, which divides by mitosis (but without a wall forming between the nuclei) and thus forms a new, coenocytic plasmodium.
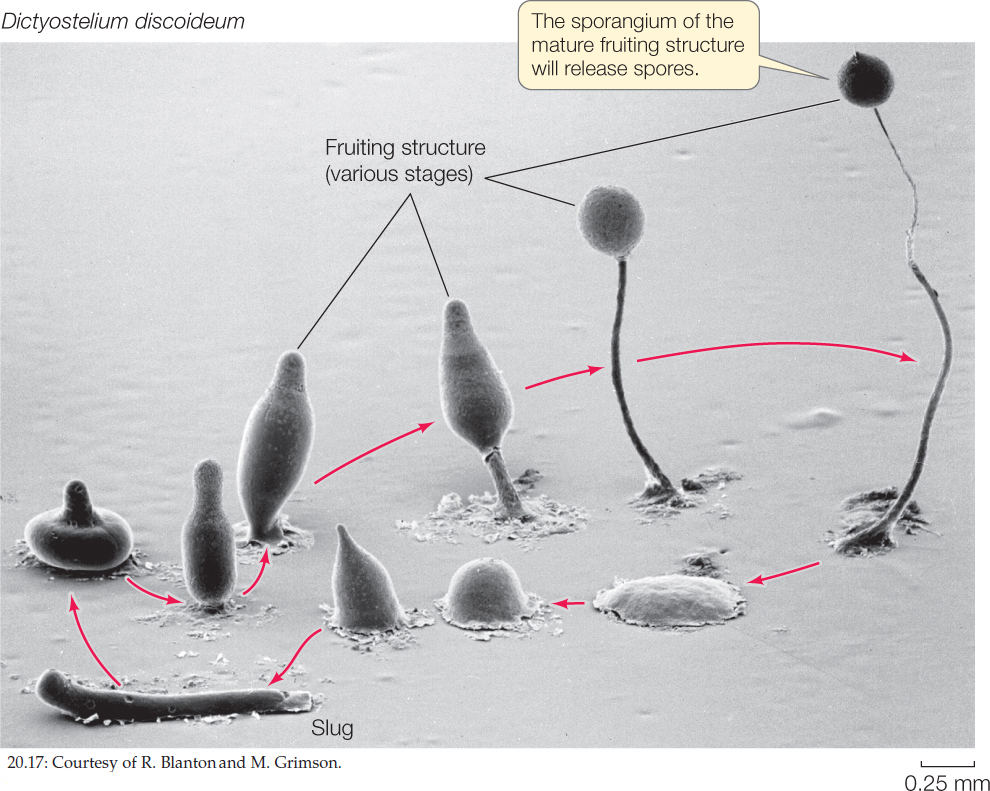
Cellular Slime Molds
Whereas the plasmodium is the basic vegetative unit of the plasmodial slime molds, an amoeboid cell is the vegetative unit of the cellular slime molds (FIGURE 20.17). Cells called myxamoebas, which have single haploid nuclei, engulf bacteria and other food particles by endocytosis and reproduce by mitosis and binary fission. This simple life cycle stage, consisting of swarms of independent, isolated cells, can persist indefinitely as long as food and moisture are available.

Go to MEDIA CLIP 20.7 Cellular Slime Mold Aggregation
PoL2e.com/mc20.7
414
When conditions become unfavorable, the cellular slime molds aggregate and form fruiting structures, as do their plasmodial counterparts. The individual myxamoebas aggregate into a mass called a slug or pseudoplasmodium. Unlike the true plasmodium of the plasmodial slime molds, however, this structure is not simply a giant sheet of cytoplasm with many nuclei. Instead, the individual myxamoebas retain their cell membranes, and therefore their identity.
A slug may migrate over a substrate for several hours before becoming motionless and reorganizing to construct a delicate, stalked fruiting structure. Cells at the top of the fruiting structure develop into thick-walled spores, which are eventually released. Later, under favorable conditions, the spores germinate, releasing myxamoebas.
The cycle from myxamoebas through slug and spores to new myxamoebas is asexual. Cellular slime molds also have a sexual cycle, in which two myxamoebas fuse. The product of this fusion develops into a spherical structure that ultimately germinates, releasing new haploid myxamoebas.
CHECKpoint CONCEPT 20.2
- Explain why the term “protists” does not refer to a formal taxonomic group.
- Contrast the major distinctive features of alveolates, excavates, stramenopiles, rhizarians, and amoebozoans. Which of these groups is most closely related to fungi and animals? What morphological evidence supports this relationship?
- The fossil record of eukaryotes in the Precambrian is poor compared with those of the Cambrian and later geological periods, even though eukaryotes were diversifying for the last billion years of the Precambrian. Can you think of some possible reasons for the better fossil record beginning in the Cambrian?
The ancient origins of major eukaryote lineages, and the adaptation of these lineages to a wide variety of lifestyles and environments, resulted in enormous protist diversity. It is not surprising, then, that reproductive modes among protists are also highly diverse.