Concept 21.2: Key Adaptations Permitted Plants to Colonize Land
How did the land plants arise? To address this question, we can compare land plants with their closest relatives among the green algae. The features that differ between the two groups include the adaptations that allowed the first land plants to survive in the terrestrial environment.
Adaptations to life on land distinguish land plants from green algae
Land plants first appeared in the terrestrial environment between 450 and 500 million years ago. How did they survive in an environment that differed so dramatically from the aquatic environment of their ancestors? While the water essential for life is everywhere in the aquatic environment, water is difficult to obtain and retain in the terrestrial environment.
424
No longer bathed in fluid, organisms on land faced potentially lethal desiccation (drying). Large terrestrial organisms had to develop ways to transport water to body parts distant from the source of the water. And whereas water provides aquatic organisms with support against gravity, a plant living on land must either have some other support system or sprawl unsupported on the ground. A land plant must also use different mechanisms for dispersing its gametes and progeny than its aquatic relatives, which can simply release them into the water.
Survival on land was facilitated by the evolution among plants of numerous adaptations, including:
- The cuticle, a coating of waxy lipids that retards water loss
- Stomata, small closable openings in leaves and stems that are used to regulate gas exchange and water loss
- Gametangia, multicellular organs that enclose plant gametes and prevent them from drying out
- Embryos, young plants contained within a protective structure
- Certain pigments that afford protection against the mutagenic ultraviolet radiation that bathes the terrestrial environment
- Thick spore walls containing a polymer that protects the spores from desiccation and resists decay
- A mutually beneficial association with fungi (mycorrhizae) that promotes nutrient uptake from the soil
The cuticle may be the most important—and the earliest—of these features. Composed of several unique waxy lipids that coat the leaves and stems of land plants, the cuticle has several functions, the most obvious and important of which is to keep water from evaporating from the plant body.
As ancient plants colonized the land, they not only adapted to the terrestrial environment, they also modified it by contributing to the formation of soil. Acids secreted by plants help break down rock, and the organic compounds produced by the breakdown of dead plants contribute nutrients to the soil. Such effects are repeated today as plants grow in new areas.
Life cycles of land plants feature alternation of generations
A universal feature of the life cycles of land plants is alternation of generations. Recall from Concept 20.3 the two hallmarks of alternation of generations:
- The life cycle includes both a multicellular diploid stage and a multicellular haploid stage.
- Gametes are produced by mitosis, not by meiosis. Meiosis produces spores that develop into multicellular haploid organisms.
If we begin looking at the land plant life cycle at the single-cell stage—the diploid zygote—then the first phase of the cycle is the formation, by mitosis and cytokinesis, of a multicellular embryo, which eventually grows into a mature diploid plant. This multicellular diploid plant is called the sporophyte (“spore plant”).
Cells contained within specialized reproductive organs of the sporophyte, called sporangia (singular sporangium), undergo meiosis to produce haploid, unicellular spores. By mitosis and cytokinesis, a spore develops into a haploid plant. This multicellular haploid plant, called the gametophyte (“gamete plant”), produces haploid gametes by mitosis. The fusion of two gametes (fertilization) forms a single diploid cell—the zygote—and the cycle is repeated (FIGURE 21.4).
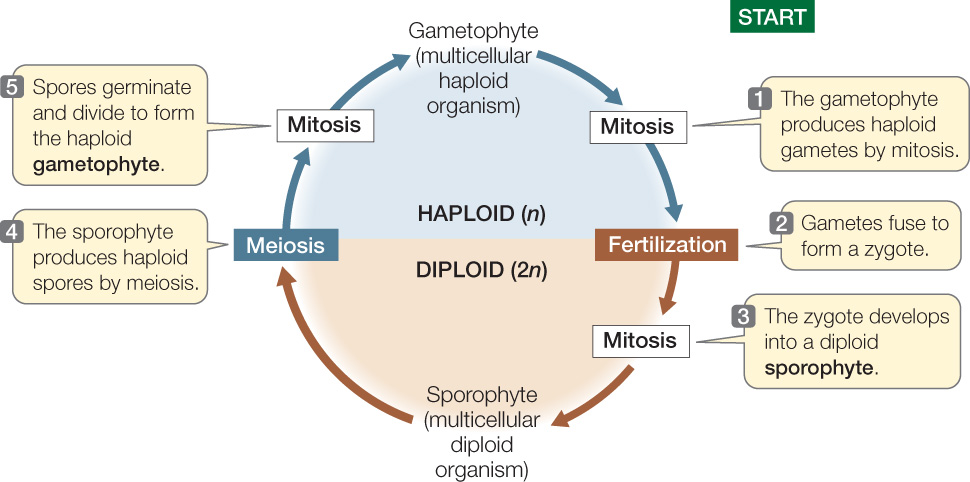
The sporophyte generation extends from the zygote through the adult multicellular diploid plant and sporangium formation. In contrast, the gametophyte generation extends from the spore through the adult multicellular haploid plant to the gametes. The transitions between the generations are accomplished by fertilization and by meiosis. In all land plants, the sporophyte and the gametophyte differ genetically: the sporophyte has diploid cells, and the gametophyte has haploid cells.
425
There is a trend toward reduction of the gametophyte generation in plant evolution. In the nonvascular land plants, the gametophyte is larger, longer-lived, and more self-sufficient than the sporophyte. In those groups that appeared later in plant evolution, however, the sporophyte is the larger, more conspicuous, longer-lived, and more self-sufficient generation.
Nonvascular land plants live where water is readily available
The living species of nonvascular land plants are the liverworts, mosses, and hornworts. These three groups are thought to be similar in many ways to the earliest land plants. Most of these plants grow in dense mats, usually in moist habitats. Even the largest of these species are only about half a meter tall, and most are only a few centimeters tall or long. Why have they not evolved to be taller? The probable answer is that they lack an efficient vascular system for transporting water and minerals from the soil to distant parts of the plant body.
The nonvascular land plants lack the true leaves, stems, and roots that characterize the vascular plants, although they have structures analogous to each. Their growth form allows water to move through the mats of plants by capillary action. They have leaflike structures that readily catch and hold any water that splashes onto them. They are small enough that minerals can be distributed throughout their bodies by diffusion. As in all land plants, layers of maternal tissue protect their embryos from desiccation. Nonvascular land plants also have a cuticle, although it is often very thin (or even absent in some species) and thus is not highly effective in retarding water loss.
Most nonvascular land plants live on the soil or on vascular plants, but some grow on bare rock, on dead and fallen tree trunks, and even on buildings. Their ability to grow on such marginal surfaces results from a mutualistic association with fungi. The earliest association of land plants with fungi dates back at least 460 million years. This mutualism probably facilitated the absorption of water and minerals, especially phosphorus, from the first soils.
LINK
Land plants of many groups have mutualistic associations with fungi, as described in Concept 22.2
Nonvascular land plants are widely distributed over six continents and even exist (albeit very locally) on the coast of the seventh, Antarctica. Most are terrestrial. Although a few species live in fresh water, these aquatic species are descended from terrestrial ones. None live in the oceans.
Liverworts
There are about 9,000 species of liverworts. Most liverworts have green, leaflike gametophytes that lie close to or flat on the ground (FIGURE 21.5A). The simplest liverworts are flat plates of cells a centimeter or so long with structures that produce sperm or eggs on their upper surfaces and rootlike filaments on their lower surfaces. The sporophyte remains attached to the larger gametophyte and rarely exceeds a few millimeters in length. Most liverworts can reproduce asexually (through simple division of the gametophyte) as well as sexually.


Go to MEDIA CLIP 21.2 Liverwort Life Cycle
PoL2e.com/mc21.2
Mosses
The most familiar of the nonvascular land plants are the mosses (FIGURE 21.5B). These hardy little plants, of which there are about 15,000 species, are found in almost every terrestrial environment. They are often found on damp, cool ground, where they form thick mats.
The mosses are the sister lineage to the vascular plants plus the hornworts (see Figure 21.1B). They share with those lineages an advance over the liverworts in their adaptation to life on land: they have stomata, which are important for both water retention and gas exchange.
Some moss gametophytes are so large that they cannot transport enough water throughout their bodies solely by diffusion. Gametophytes and sporophytes of many mosses contain a type of cell called a hydroid, which dies and leaves a tiny channel through which water can travel. The hydroid is functionally similar to the tracheid, the characteristic water-conducting cell of vascular plants, but it lacks the lignin and the cell-wall structure that characterize tracheids. The possession of hydroids shows that the term “nonvascular land plant” is somewhat misleading when applied to mosses. Despite their simple systems of internal transport, however, the mosses are not considered vascular plants because they lack tracheids and other vascular tissues.
426
Hornworts
The approximately 100 species of hornworts are so named because their sporophytes often look like little horns (FIGURE 21.5C). Hornworts have two characteristics that distinguish them from liverworts and mosses. First, the cells of hornworts each contain a single large, platelike chloroplast, whereas the cells of the other two groups contain numerous small, lens-shaped chloroplasts. Second, of the sporophytes in all three groups, those of the hornworts come closest to being capable of growth without a set limit. Liverwort and moss sporophytes have a stalk that stops growing as the spore-producing structure matures, so elongation of the sporophyte is strictly limited. The hornwort sporophyte, however, has no stalk, and it is persistently green (a trait shared with vascular plants). A basal region of the sporangium remains capable of indefinite cell division, continuously producing new spore-bearing tissue above. The sporophytes of some hornworts growing in mild and continuously moist conditions can become as tall as 20 centimeters. Eventually, however, the sporophyte’s growth is limited by the lack of a transport system.
Hornworts have evolved a symbiotic relationship that promotes their growth by providing them with greater access to nitrogen, which is often a limiting resource. Hornworts have internal cavities filled with mucilage, and the cavities are often populated by symbiotic cyanobacteria that convert atmospheric nitrogen gas into a form of nitrogen usable by their host plant.
The sporophytes of nonvascular land plants are dependent on the gametophytes
In the nonvascular land plants, the conspicuous green structure visible to the naked eye is the gametophyte. The gametophyte is photosynthetic and is therefore nutritionally independent. The sporophyte may or may not be photosynthetic, but it is always nutritionally dependent on the gametophyte and remains permanently attached to it.
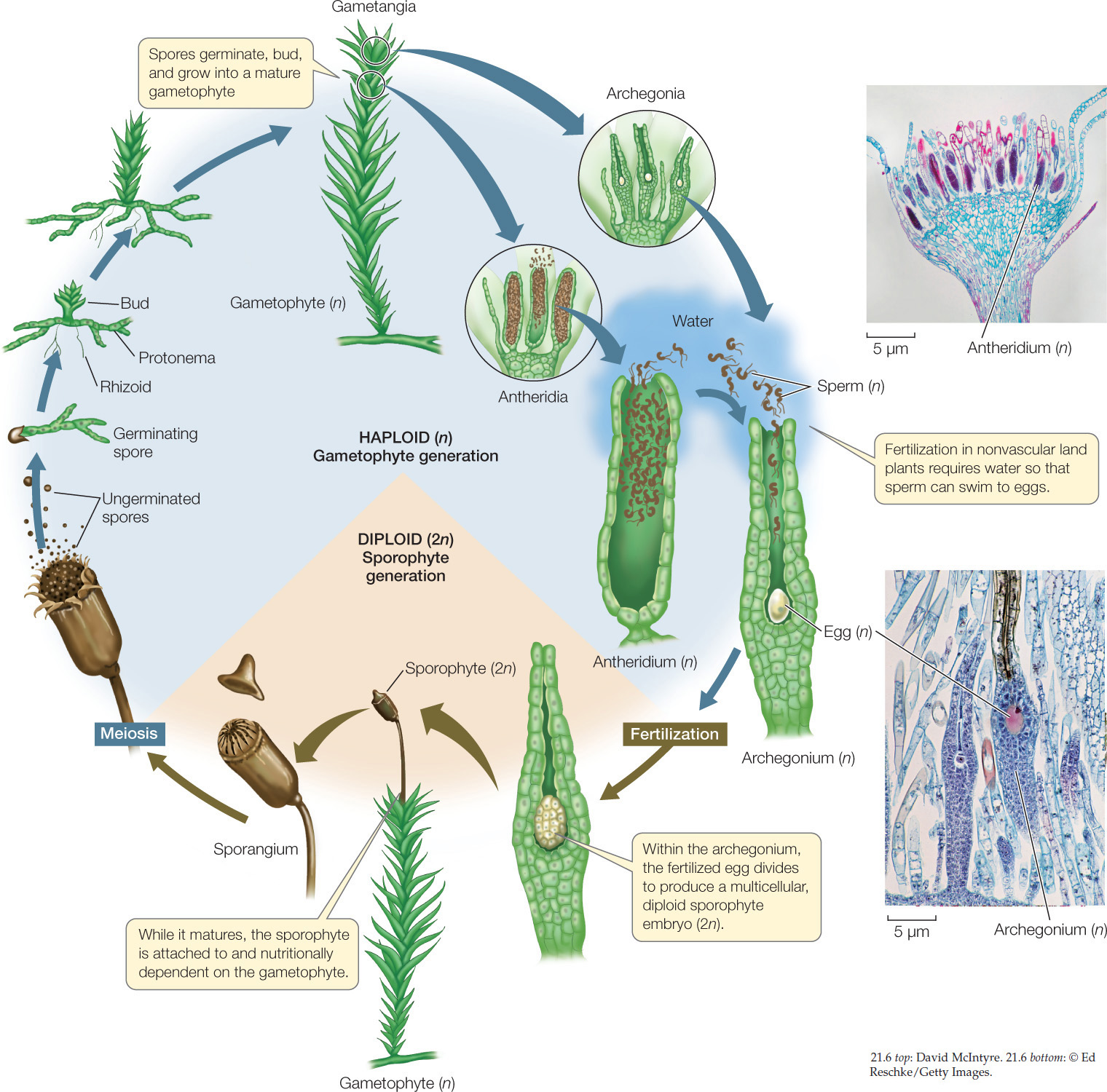
FIGURE 21.6 illustrates a moss life cycle that is typical of the life cycles of nonvascular land plants. A sporophyte produces unicellular haploid spores as products of meiosis within a sporangium. When a spore germinates, it gives rise to a multicellular haploid gametophyte whose cells contain chloroplasts and are thus photosynthetic. Eventually gametes form within specialized sex organs, called the gametangia. The archegonium is a multicellular, flask-shaped female sex organ that produces a single egg. The antheridium is a male sex organ in which sperm, each bearing two flagella, are produced in large numbers. Both archegonia and antheridia are produced on the same individual, so each individual has both male and female reproductive structures. Adjacent individuals often fertilize one another’s gametes, however, which helps maintain genetic diversity in the population.

Go to ANIMATED TUTORIAL 21.1 Life Cycle of a Moss
PoL2e.com/at21.1

Go to MEDIA CLIP 21.3 Bryophyte Reproduction
PoL2e.com/mc21.3
427
Once released from the antheridium, the sperm must swim or be splashed by raindrops to a nearby archegonium on the same or a neighboring plant—a constraint that reflects the aquatic origins of the nonvascular land plants’ ancestors. The sperm are aided on their journey by chemical attractants released by the egg or the archegonium. Before sperm can enter the archegonium, however, certain cells in the neck of the archegonium must break down, leaving a water-filled canal through which the sperm can swim to complete their journey. Notice that all of these events require liquid water.
428
Once sperm arrive at an egg, the nucleus of a sperm fuses with the egg nucleus to form a diploid zygote. Mitotic divisions of the zygote produce a multicellular, diploid sporophyte embryo. The sporophyte matures and produces a sporangium, within which meiotic divisions produce spores and thus the next gametophyte generation.
CHECKpoint CONCEPT 21.2
- Describe several adaptations of plants to the terrestrial environment, and describe the distribution of those adaptations in liverworts, mosses, and hornworts.
- Explain what is meant by alternation of generations.
- What aspects of the reproductive cycle of nonvascular land plants appear to have been retained from their aquatic ancestors?
Further adaptations to the terrestrial environment appeared as plants continued to evolve. One of the most important of these later adaptations was the appearance of vascular tissues—the characteristic that defines the vascular plants.