Nature and Nurture
Everything about you—from your physical structure, intellectual capacity, and personality characteristics to your preferences in hobbies and food—is a joint consequence of the interaction between the genetic material you inherited from your parents and the environments you have experienced from conception to the present moment. These two factors—heredity and environment—work in concert to influence both the ways in which you are like other people and the ways in which you are unique.
Long before there was any understanding of the principles of heredity, people were aware that some traits and characteristics “run in families” and that this tendency was somehow related to procreation. For as long as there have been domesticated animals, for example, farmers have practiced selective breeding to improve certain characteristics of their livestock, such as the size of their horses and the milk yield of their goats, cows, or yaks. People have also long been aware that the environment plays a role in development—that a nutritious diet, for example, is necessary for livestock to produce a good milk supply or fine-quality wool. When scientists first began to investigate the contributions of heredity and environment to development, they generally emphasized one factor or the other as the prime influence—heredity or environment, nature or nurture. In nineteenth-century England, for example, Francis Galton (1869/1962), a cousin of Charles Darwin, identified men who had achieved “eminence” in a variety of fields and concluded that talent runs in families, because very close relatives of an eminent man (his father, brother, son) were more likely to be high achievers themselves than were less close relatives.
Among Galton’s cases of closely related eminent men were John Stuart Mill and his father, both respected English philosophers. However, Mill himself pointed out that most of Galton’s eminent men were members of well-to-do families. In his view, the relation between the achievement of these eminent men and their kinship had less to do with biological ties than with the fact that they were similar in economic well-being, social status, education, and other advantages and opportunities. In short, according to Mill, Galton’s subjects rose to eminence more because of environmental factors than hereditary ones.

88
Our modern understanding of how characteristics are transmitted from parent to offspring originated with insights achieved by Gregor Mendel, a nineteenth-century Austrian monk who observed distinct patterns of inheritance in the pea plants that he cross-bred in his monastery garden. Some aspects of these inheritance patterns were later discovered to occur in all living things. A much deeper understanding of how genetic influences operate came with James Watson and Francis Crick’s 1953 identification of the structure of DNA, the basic component of hereditary transmission.
genome the complete set of genes of any organism
Since that landmark discovery, enormous progress has been made in deciphering the genetic code. Researchers have mapped the entire genome—the complete set of genes—of myriad species of plants and animals, including chickens, mice, chimpanzees, and humans, and even several extinct species, including our closest evolutionary relative, Neanderthals (R. E. Green et al., 2010). In 2010, a consortium of geneticists began working to sequence the genomes of 10,000 vertebrate species (Lander, 2011), the expectation being that examining the genomes of such a diverse set of species will provide knowledge not only about those species but also about human evolution and the way genes function. Comparisons of the genomes of various species have already revealed much about our human genetic endowment, and they have provided numerous surprises.

One surprise was the number of genes that humans have: the current estimate of around 21,000 genes is far fewer than previous estimates, which ranged from 35,000 to more than 100,000 genes (Clamp et al., 2007). A second major surprise was that most of those genes are possessed by all living things. We humans share a large proportion of our genes with bears, barnacles, beans, and bacteria. Most of our genes are devoted, in decreasing order, to making us animals, vertebrates, mammals, primates, and—finally—humans. In the next section, we will look at a third surprise, one that may turn out to be a blockbuster.
As researchers have achieved better understanding of the role of hereditary factors in development, they have also come to appreciate the limits of what these factors can account for on their own. Similarly, as knowledge has grown concerning the influence of experience on development, it has become clear that experience alone rarely provides a satisfactory account. Development results from the close and continual interplay of nature and nurture—of genes and experience—and this interplay is the focus of the following section.
Genetic and Environmental Forces
genotype the genetic material an individual inherits
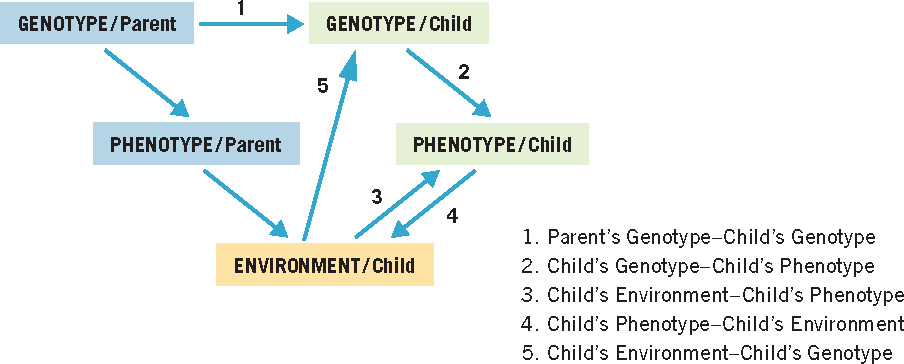
The interplay of genes and experience is exceedingly complex. To simplify our discussion of interactions among genetic and environmental factors, we will organize it around the model of hereditary and environmental influences shown in Figure 3.1. Three key elements of the model are the genotype—the genetic material an individual inherits; the phenotype—the observable expression of the genotype, including both body characteristics and behavior; and the environment—every aspect of the individual and his or her surroundings (including prenatal experience) other than the genes themselves.
phenotype the observable expression of the genotype, including both body characteristics and behavior
89
environment every aspect of an individual and his or her surroundings other than genes
These three elements are involved in five relations that are fundamental in the development of every child: (1) the parents’ genetic contribution to the child’s genotype; (2) the contribution of the child’s genotype to his or her own phenotype; (3) the contribution of the child’s environment to his or her phenotype; (4) the influence of the child’s phenotype on his or her environment; and (5) the influence of the child’s environment on his or her genotype. We will now consider each of these relations in turn.
1. Parent’s Genotype–Child’s Genotype
chromosomes molecules of DNA that transmit genetic information; chromosomes are made up of DNA
DNA (deoxyribonucleic acid) molecules that carry all the biochemical instructions involved in the formation and functioning of an organism
Relation 1 involves the transmission of genetic material—chromosomes and genes—from parent to offspring. You caught a glimpse of this process in Chapter 2, when we discussed the gametes (one from the mother and one from the father) that conjoin at conception to create a zygote. The nucleus of every cell in the body contains chromosomes, long threadlike molecules made up of two twisted strands of DNA (deoxyribonucleic acid). DNA carries all the biochemical instructions involved in the formation and functioning of an organism. These instructions are “packaged” in genes, the basic unit of heredity in all living things. Genes are sections of chromosomes. More specifically, each gene is a segment of DNA that is the code for the production of particular proteins. Some proteins are the building blocks of the body’s cells; others regulate the cells’ functioning. Genes affect development and behavior only through the manufacture of proteins: “DNA’s information translated into flesh and blood” (J. S. Levine & Suzuki, 1993, p. 19).
genes sections of chromosomes that are the basic unit of heredity in all living things
But here is the blockbuster surprise we mentioned earlier: researchers have discovered that genes—at least “genes” as they have been traditionally defined—make up only about 2% of the human genome (Mouse Genome Sequencing Consortium, 2002). Much of the rest of our genome—once thought to be “junk” DNA—turns out to play a supporting role in influencing genetic transmission by regulating the activity of protein-coding genes (e.g., Mendes Soares & Valcárcel, 2006). Just how much of this noncoding DNA is vital to functioning, and precisely how it works, is, as of today, shrouded in mystery and controversy. Given the pace of genetic research, however, tomorrow may be a different story.
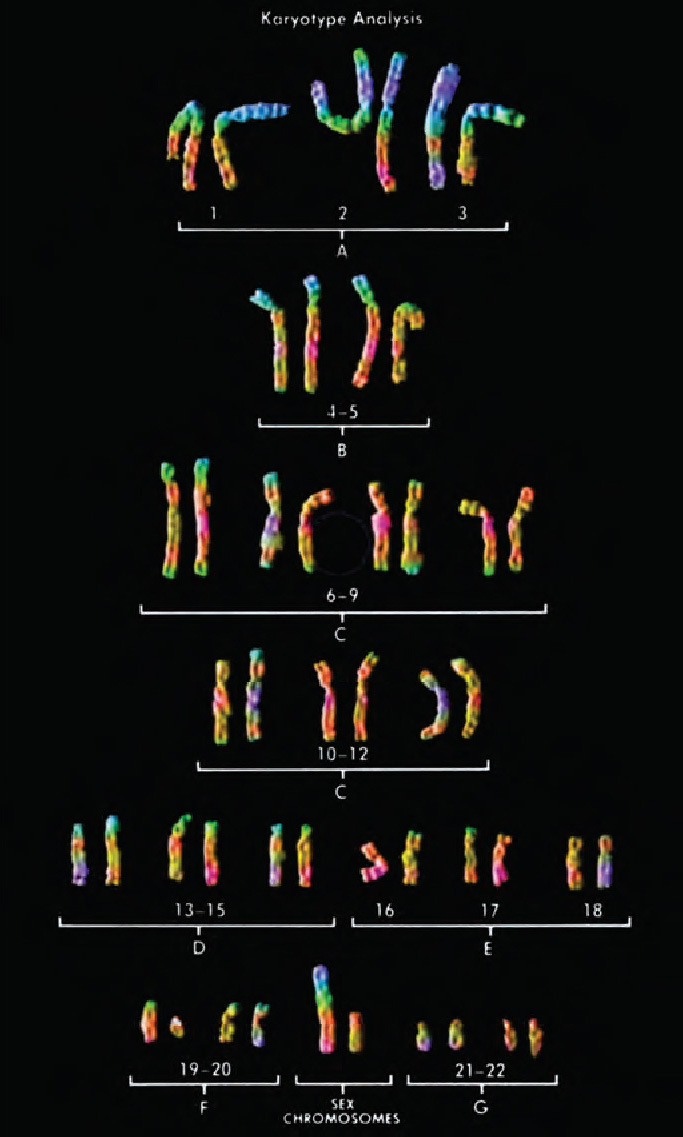
Human heredity Humans normally have a total of 46 chromosomes in the nucleus of each cell, except egg and sperm cells. (Recall from Chapter 2 that, as a result of the type of cell division that produces gametes, eggs and sperm each contain only 23 chromosomes.) These 46 chromosomes are actually 23 pairs (Figure 3.2). With one exception—the sex chromosomes—the two members of each chromosome pair are of the same general size and shape (roughly the shape of the letter X). Furthermore, each chromosome pair carries, usually at corresponding locations, genes of the same type—that is, sequences of DNA that are relevant to the same traits. One member of each chromosome pair was inherited from each parent. Thus, every individual has two copies of each gene, one on the chromosome inherited from the father and one on the chromosome from the mother. Your biological children will each receive half of your genes, and your grandchildren will have one-quarter (just as you have half your genes in common with each of your biological parents and one-fourth with each grandparent).
90
sex chromosomes the chromosomes (X and Y) that determine an individual’s gender
Sex determination As noted, the sex chromosomes, which determine an individual’s sex, are an exception to the general pattern of chromosome pairs being the same size and shape and carrying corresponding genes. Females have two identical, largish sex chromosomes, called X chromosomes, but males have one X chromosome and one much smaller Y chromosome (so called because it has the shape of the letter Y). Because a female has only X chromosomes, the division of her germ cells results in all her eggs having an X. However, because a male is XY, half his sperm contain an X chromosome and half contain a Y. For this reason, it is always the father who determines the sex of offspring: if an X-bearing sperm fertilizes an egg, a female (XX) zygote results; if an egg is fertilized by a Y-bearing sperm, the zygote is male (XY). It is the presence of a Y chromosome—not the fact of having only one X chromosome—that makes an individual male. A gene on the Y chromosome encodes the protein that triggers the prenatal formation of testes by activating genes on other chromosomes. Subsequently, the testes produce the hormone testosterone, which takes over the molding of maleness (Jegalian & Lahn, 2001).
Diversity and individuality As we have noted, genes guarantee that humans will be similar to one another in certain ways, both at the species level (we are all bipedal and have opposable thumbs, for example) and at the individual level (i.e., family resemblances). Genes also guarantee differences at both levels. Several mechanisms contribute to genetic diversity among people.

mutation a change in a section of DNA
One such mechanism is mutation, a change that occurs in a section of DNA. Some mutations are random, spontaneous errors; others are caused by environmental factors. Most are harmful. Those that occur in germ cells can be passed on to offspring; many inherited diseases and disorders originate from a mutated gene. (Box 3.1 discusses the genetic transmission of diseases and disorders.)
Occasionally, however, a mutation that occurs in a germ cell or early in prenatal development makes individuals more viable, that is, more likely to survive—perhaps by increasing their resistance to some disease or by increasing their ability to adapt to some crucial aspect of their environment. Such mutations provide the basis for evolution. This is because a person with the favorable mutated gene is more likely to survive long enough to produce offspring, who, in turn, are likely to possess the mutated gene, thus heightening their own chance of surviving and reproducing. Across generations, these favorable genes proliferate in the gene pool of the species.
91
crossing over the process by which sections of DNA switch from one chromosome to the other; crossing over promotes variability among individuals
A second mechanism that promotes variability among individuals is the random assortment of chromosomes in the formation of egg and sperm. During germ-cell division, the 23 pairs of chromosomes are shuffled randomly, with chance determining which member of each pair goes into each new egg or sperm. This means that, for each germ cell, there are 223, or 8.4 million, possible combinations of chromosomes. Thus, when a sperm and an egg unite, the odds are essentially zero that any two individuals—even members of the same family—would have the same genotype (except, of course, identical twins). Further variation is introduced by the fact that when germ cells divide, the two members of a pair of chromosomes sometimes swap sections of DNA. As a result of this process, referred to as crossing over, some of the chromosomes that parents pass on to their offspring are constituted differently from their own.
2. Child’s Genotype–Child’s Phenotype
We now turn to Relation 2 in Figure 3.1, the relation between one’s genotype and one’s phenotype. Our examination of the genetic contribution to the phenotype begins with a key fact: although every cell in your body contains copies of all the genes you received from your parents, only some of those genes are expressed. At any given time in any cell in the body, some genes are active (turned on), while others are not. Some genes that are hard at work in neurons, for example, are totally at rest in toenail cells. As you will see, there are several reasons for this.
Gene expression: Developmental changes Genes influence development and behavior only when they are turned on, and human development proceeds normally, from conception to death, only if genes get switched on and off in the right place, at the right time, and for the right length of time. Some genes are turned on in only a few cells and for only a few hours and then are switched off permanently. This pattern is typical during embryological development when, for example, the genes that are turned on in certain cells lead them to specialize for arm, hand, and fingerprint formation. Other genes are involved in the basic functioning of almost all cells almost all the time.
regulator genes genes that control the activity of other genes
The switching on and off of genes is controlled primarily by regulator genes. The activation or inactivation of one gene is always part of a chain of genetic events. When one gene is switched on, it causes another gene to turn on or off, which has an impact on the status of yet other genes. Thus, genes never function in isolation. Instead, they belong to extensive networks in which the expression of one gene is a precondition for the expression of another, and so on. The continuous switching on and off of genes underlies development throughout life, from the initial prenatal differentiation of cells to the gene-induced events of puberty to many of the changes related to aging.
External factors can affect the switching on and off of genes. A dramatic example is the effect of thalidomide on limb development (described in Chapter 2), in which the sedative interferes with the functioning of genes underpinning normal growth factors (Ito et al., 2010). Another example comes from the fact that early visual experience is necessary for the normal development of the visual system, because it causes the switching on of certain genes, which, in turn, switch on other genes in the visual cortex (Maya-Vetencourt & Origlia, 2012). The ramifications of decreased visual experience are observed in cases of children with cataracts that are not removed early in life, as discussed later in this chapter.
92
The fact that regulator genes can repeatedly switch other genes on and off in different patterns means that a given gene can function multiple times in multiple places during development. All that is required is that the gene’s expression be controlled by different regulator genes at different times. This on-again, off-again functioning of individual genes results in enormous diversity in genetic expression. By analogy, consider the fact that this book is written with only 26 letters and probably only a few thousand different words made up of combinations of those letters. The meaning comes from the order in which the letters occur, the order in which they have been “switched on and off” by the authors.
alleles two or more different forms of a gene
Gene expression: Dominance patterns Many of an individual’s genes are never expressed; some others are only partially expressed. One reason for this is the fact that about one-third of human genes have two or more different forms, known as alleles. The alleles of a given gene influence the same trait or characteristic (e.g., eye color), but they contribute to different developmental outcomes (e.g., brown, blue, hazel, gray eyes).
dominant allele the allele that, if present, gets expressed
recessive allele the allele that is not expressed if a dominant allele is present
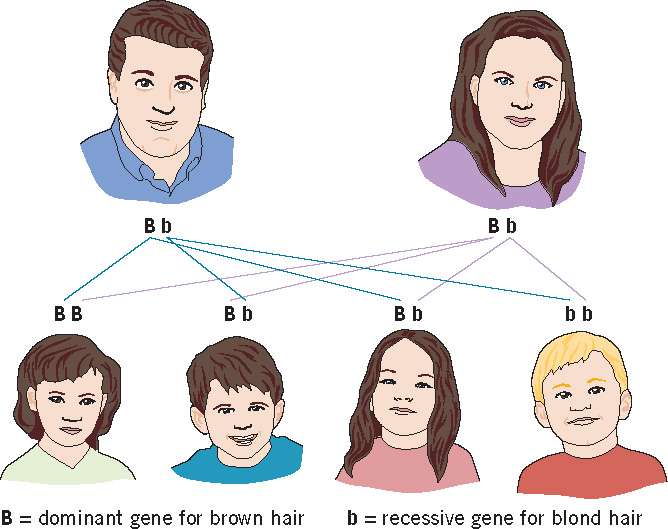
Let’s consider the simplest pattern of gene expression—the one discovered by Mendel and referred to as the dominant–recessive pattern. The explanation for this pattern (unknown to Mendel) is that some genes have only two alleles, one of which is dominant and the other recessive. In this pattern, there are two possibilities: (1) a person can inherit two of the same allele—two dominant or two recessive—and thus be homozygous for the trait in question; or (2) the person can inherit two different alleles—one dominant and the other recessive—and thus be heterozygous for the trait. When an individual is homozygous, with either two dominant or two recessive alleles, the corresponding trait will be expressed. When an individual is heterozygous for a trait, the instructions of the dominant allele will be expressed (see Figure 3.3).
homozygous having two of the same allele for a trait
heterozygous having two different alleles for a trait
To illustrate, let us consider two traits of no importance to human survival: the ability to roll one’s tongue lengthwise and curliness of hair. If you can roll your tongue lengthwise into the shape of a tube, then at least one, but not necessarily both, of your parents must also possess this remarkable but useless talent. From this statement (and Figure 3.3), you should be able to figure out that tongue rolling is governed by a dominant allele. In contrast, if you have straight hair, then both of your parents must carry an allele for this trait, although it is possible that neither of them actually has straight hair. This is because straight hair is governed by a recessive gene, and curly hair is governed by a dominant gene.
The sex chromosomes present an interesting wrinkle in the story of dominance patterns. The X chromosome carries roughly 1500 genes, whereas the much smaller Y chromosome carries only about 200. Thus, when a female inherits a recessive allele on the X chromosome from her mother, she is likely to have a dominant allele on the chromosome from her father to suppress it, so she will not express the trait in question. In contrast, when a male inherits the same recessive allele on the X chromosome from his mother, he likely will not have a dominant allele from his father to override it, so he will express the trait. This difference in sex-linked inheritance is one reason for the greater vulnerability of males described in Chapter 2 (Box 2.2): they are more likely to suffer a variety of inherited disorders caused by recessive alleles on their X chromosome (see also Box 3.1).
93
Despite the traditional emphasis given to it, the dominant–recessive pattern of inheritance, in which a single gene affects a particular trait, pertains to relatively few human traits—such as hair color, blood type, abundance of body hair, and the like—as well as to a large number of genetic disorders Much more commonly, a single gene can affect multiple traits; both alleles can be fully expressed or blended in heterozygous individuals; and some genes are expressed differently, depending on whether they are inherited from the mother or from the father.
polygenic inheritance inheritance in which traits are governed by more than one gene
Inheritance patterns are vastly more complicated for most of the traits and behaviors that are of primary interest to behavioral scientists. These traits, such as shyness, aggression, thrill-seeking, and language learning, involve polygenic inheritance, in which several different genes contribute to any given phenotypic outcome. Gene-outcome linkages are particularly difficult to detect when many genes are involved. For this reason, you should be skeptical whenever you encounter newspaper headlines announcing the discovery of “a gene for” a complex human trait or predisposition.
3. Child’s Environment–Child’s Phenotype
We now come to Relation 3 in our model—the impact of the environment on the child’s phenotype. (Remember, the environment includes everything not in the genetic material itself, including the variety of prenatal experiences discussed in Chapter 2.) As the model indicates, the child’s observable characteristics result from the interaction between environmental factors and the child’s genetic makeup.
norm of reaction all the phenotypes that can theoretically result from a given genotype in relation to all the environments in which it can survive and develop
Because of the continuous interaction of genotype and environment, a given genotype will develop differently in different environments. This idea is expressed by the concept of the norm of reaction (Dobzhansky, 1955), which refers to all the phenotypes that could theoretically result from a given genotype in relation to all the environments in which it could survive and develop. According to this concept, for any given genotype developing in varying environments, a range of outcomes would be possible. A child with a given genotype would probably develop quite differently in a loving, supportive family than he or she would in an alienating, abusive family. (Figure 3.4 offers a classic illustration of the norm of reaction in a genotype–environment interaction.)
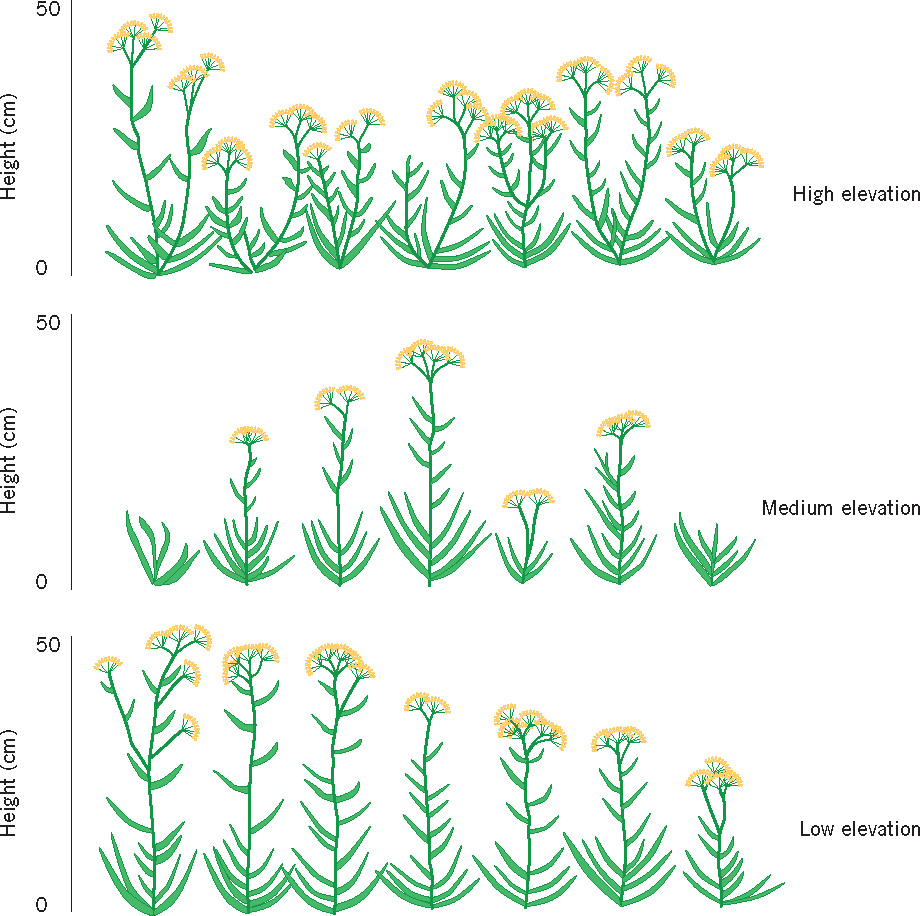
Examples of genotype–environment interaction Genotype–environment interactions can be studied directly by randomly assigning nonhuman animals with known genotypes to be raised in a wide variety of environmental conditions. If genetically identical animals develop differently in different environments, researchers can infer that environmental factors must be responsible for the different developmental outcomes. Scientists cannot, of course, randomly assign humans to different rearing conditions, but there are powerful naturally occurring examples of genotype–environment interactions for humans.
phenylketonuria (PKU) a disorder related to a defective recessive gene on chromosome 12 that prevents metabolism of phenylalanine
One such example is phenylketonuria (PKU), a disorder related to a defective recessive gene on chromosome 12. Individuals who inherit this gene from both parents cannot metabolize phenylalanine, an amino acid present in many foods (especially red meats) and in artificial sweeteners. If they eat a normal diet, phenylalanine accumulates in the bloodstream, causing impaired brain development that results in severe intellectual impairment. However, if infants with the PKU gene are identified shortly after birth and placed on a stringent diet free of phenylalanine, intellectual impairment can be avoided, as long as the diet is carefully maintained. Thus, a given genotype results in quite different phenotypes—cognitive disability or relatively normal intelligence—depending on environmental circumstances. Because early detection of this genetic disorder has such a positive effect on children’s developmental outcomes, all newborn infants in the United States are routinely screened for PKU, as well as for a number of other severe and easily detected genetic disorders.
94
Box 3.1: applications: Genetic Transmission of Disorders
Thousands of human disorders—many of them extremely rare—are presently known to have genetic origins. Although our discussion here will focus on the behaviors and psychological symptoms associated with such disorders, most of them also involve a variety of physical symptoms, often including unusual physical appearance (e.g., distorted facial features), organ defects (e.g., heart problems), and atypical brain development. These and other genetically based conditions can be inherited in several different ways.
Dominant–Recessive Patterns
Many genetic disorders involve the dominant–recessive pattern of inheritance, occurring only when an individual has two recessive alleles for the condition. To date, more than 2850 such disorders have been identified (Lander, 2011). Recessive-gene disorders include PKU (discussed on pages 94 and 96) and sickle-cell anemia (discussed below), as well as Tay-Sachs disease, cystic fibrosis, and many others. Disorders that are caused by a dominant gene include Huntington disease (a progressive and always fatal degenerative condition of the brain) and neurofibromatosis (a disorder in which nerve fibers develop tumors). A combination of severe speech, language, and motor difficulties that is common in a particular family in England has been traced to a mutation of a single gene (referred to as FOXP2) that acts in a dominant fashion (see S. E. Fisher & Scharff, 2009).
In some cases, a single gene can have both harmful and beneficial effects. One such case is sickle-cell disease, in which red blood cells are sickle-shaped rather than round, diminishing their capacity to transport oxygen. This disease, which can be debilitating and sometimes fatal, affects about 1 of every 500 African Americans. It is a recessive-gene disorder, so individuals who are homozygous for this trait (inheriting two sickle-cell genes, one from each parent) will suffer from the disease. Individuals who are heterozygous for this trait (carrying one normal and one sickle-cell gene) have some abnormality in their blood cells but usually experience no negative effects. In fact, if they live in regions of the world—like West Africa—where malaria is common, they benefit, because the sickle cells in their blood confer resistance against this deadly disease. In nineteenth-century Africa, malaria came to be known as the “White man’s disease” because so many European explorers, lacking the sickle-cell gene, died of it.
Note that even when the root cause of a disorder is a single gene, it does not mean that that one gene is responsible for all manifestations of the disorder. The single gene simply starts a cascade of events, turning on and turning off multiple genes with effects on many different aspects of the individual’s subsequent development.
Polygenic Inheritance
Many common human disorders are believed to result from interactions among multiple inherited genes, often in conjunction with environmental factors. Among the many diseases in this category are some forms of cancer and heart disease, Type 1 and Type 2 diabetes, and asthma. Psychiatric disorders, such as schizophrenia, and behavior disorders, such as attention-deficit hyperactivity disorder, probably also involve multiple genes. More than 1100 gene loci affecting common traits and diseases have been identified to date, due largely to continually improving methods for genetic epidemiology (Lander, 2011).
Sex-Linked Inheritance
As mentioned in the text, some single-gene conditions are carried on the X chromosome and are much more common in males. (Females can inherit such conditions, but only if they inherit the culprit recessive alleles on both of their X chromosomes.) Sex-linked disorders range from relatively minor problems, like male-pattern baldness and red–green color blindness, to very serious problems, including hemophilia and Duchenne muscular dystrophy. Another sex-linked disorder is fragile-X syndrome, which involves mutations in the X chromosome and is the most common inherited form of intellectual disability.
Chromosomal Anomalies
Some genetic disorders originate with errors in germ-cell division that result in a zygote that has either more or less than the normal complement of chromosomes. Most such zygotes cannot survive, but some do. Down syndrome most commonly originates when the mother’s egg cells do not divide properly, and an egg that is fertilized contains an extra copy of chromosome 21. The probability of such errors in cell division increases with age, with the incidence of giving birth to a child with Down syndrome being markedly higher for women older than 35. (Increased paternal age has also been linked to the incidence of Down syndrome, though to a lesser extent [De Souza, Alberman, & Morris, 2009; Hurles, 2012]). The boy pictured below shows some of the facial features common to individuals with Down syndrome, which is also marked by intellectual disability (ranging from mild to severe), a number of physical problems, and a sweet temperament.
Other genetic disorders arise from extra or missing sex chromosomes. For example, Klinefelter syndrome, which affects between 1 in 500 to 1000 males in the United States, involves an extra X chromosome (XXY). The physical signs of this syndrome, which can include small testes and elongated limbs, often go unnoticed, but infertility is common. Turner syndrome, which affects 1 in 2500 U.S. women, involves a missing X chromosome (XO) and is usually characterized by short stature, stunted sexual development at puberty, and infertility.
Gene Anomalies
Just as genetic disorders can originate from extra or missing chromosomes, so too can they result from extra, missing, or abnormal genes. One intriguing instance is Williams syndrome. This rare genetic disorder involves a variety of cognitive impairments, most noticeably in spatial and visual skills, but relatively less impairment in language ability (e.g., Musolino & Landau, 2012; Skwerer & Tager-Flusberg, 2011). Individuals with Williams syndrome are also typically characterized by outgoing personalities and friendliness paired with anxiety and phobias. This condition has been traced to the deletion of a small section of approximately 25 genes on chromosome 7. Some individuals, however, have a smaller deletion; in these cases, the degree of impairment is decreased, suggesting a clear relationship between the number of genes deleted and the resulting phenotype (Karmiloff-Smith et al., 2012). Interestingly, some individuals show a duplication of the same section of genes that is deleted in Williams syndrome. In this disorder, known as 7q11.23 duplication syndrome, the pattern of abilities and disabilities is flipped, with individuals exhibiting relatively weak speech and language abilities paired with relatively strong visuo-spatial skills (Mervis & Velleman, 2011; Osborne & Mervis, 2007).
95
Regulator Gene Defects
Many disorders are thought to originate from defects in regulator genes, which control the expression of other genes. For example, a defect in the regulator gene that initiates the development of a male can interrupt the normal chain of events, occasionally resulting in a newborn who has female genitalia but is genetically male. Such cases often come to light when a young woman fails to begin menstruating or when a fertility clinic discovers that the reason a couple has failed to conceive is that the person trying to get pregnant is genetically male.

Unidentified Genetic Basis
In addition to the known gene-disorder links, there are many syndromes whose genetic origins are clear from their inheritance patterns but whose specific genetic cause has yet to be identified. For example, dyslexia is a highly heritable reading disability that probably stems from a variety of gene-based conditions. Another example is Tourette syndrome. Individuals with this disorder generally display a variety of tics, ranging from involuntary twitching and jerking to compulsively blurting out obscenities. Research suggests that Tourette syndrome probably involves a complex pattern of inheritance, making precise determination of the cause very difficult (O’Rourke et al., 2009).
The same is true for autism spectrum disorder (ASD), which includes both autism and Asperger syndrome and involves a wide range of deficits in social skills and communication. In 2008, in various U.S. districts monitored by the Centers for Disease Control, it was estimated that the ASD prevalence rate among 8-year-olds was 1 in 88 children (age 8 is thought to be the age of peak prevalence), with boys being 5 times more likely than girls to be identified as having the disorder (Baio, 2012). The diagnosis of ASD is based on major impairments in social interaction and communication skills and a limited set of interests or repetitive behaviors. Individuals with Asperger syndrome tend to have a milder array of symptoms and usually do not experience difficulties in language development.
ASD includes individuals with not only a range of disabilities but also, in some cases, remarkable talents in a narrowly focused area, such as mathematics or drawing. ASD is known to be highly heritable: twin studies have revealed that identical twins (who share 100% of their genes) are more than twice as likely as fraternal twins (who share 50% of their genes) to share an autism diagnosis (Ronald & Hoekstra, 2011). The difficulty in identifying the specific genetic basis for autism spectrum disorder is highlighted by the fact that, at present, there are more than 100 candidate genes associated with ASD (Geschwind, 2011; L. M. Xu et al., 2012).
The number of children diagnosed with ASD has increased dramatically in recent years. Indeed, in the districts tracked by the Centers for Disease Control, the ASD prevalence estimates for 8-year-olds in 2008 represented a 78% increase over those for 2002 (Baio, 2012). Part of the increase is believed to be due to greater public awareness of the syndrome, leading to a higher level of detection by parents, teachers, and doctors. In addition, current diagnostic criteria are broader than those of the past. It is thus unclear to what degree the increased level of diagnoses accurately reflects a change in the actual incidence of ASD (e.g., Gernsbacher, Dawson, & Goldsmith, 2005).
One factor that was highly publicized as a possible cause of the so-called autism epidemic—the MMR vaccine that is routinely given to young children to prevent measles, mumps, and rubella—has been definitively ruled out (A. W. McMahon et al., 2008; Price et al., 2010). Indeed, the original study reporting a link between the MMR vaccine and ASD (Wakefield et al., 1998) has been shown to be fraudulent and has been retracted (Godlee, Smith, & Marcovitch, 2011). Unfortunately, however, some parents continue to deny their children this important vaccine, needlessly putting them at risk for the illnesses that the vaccine prevents.
96
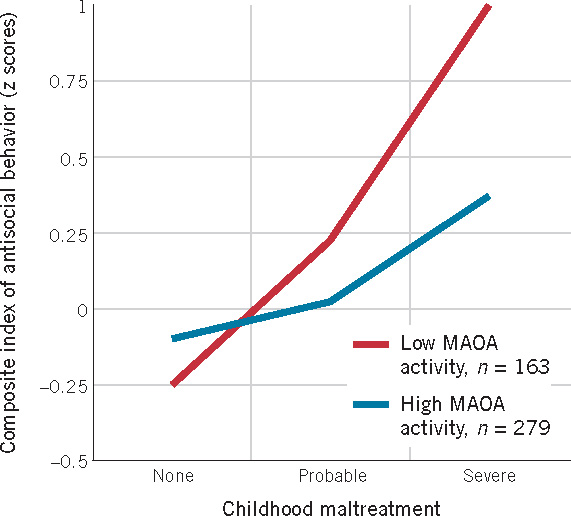
A second example of a genotype–environment interaction comes from an important study showing that the effects of abusive parenting vary in severity as a function of the child’s genotype (Caspi et al., 2002). The researchers wanted to determine why some children who experience severe maltreatment become violent and antisocial as adults, whereas others who are exposed to the same abuse do not. The results, shown in Figure 3.5, revealed the importance of a combination of environmental and genetic factors leading to antisocial outcomes—suffering abusive treatment as a child and possessing a particular variant of MAOA, an X-linked gene known to inhibit brain chemicals associated with aggression. Young men who had a relatively inactive version of the MAOA gene, and who had experienced severe maltreatment, grew up to be more antisocial than other men. More concretely, 85% of the maltreated group with the relatively inactive gene developed some form of antisocial behavior, and they were almost 10 times more likely to be convicted of a violent crime. The important point here is that neither factor by itself (possessing the inactive MAOA gene or being abused) predisposed boys to become highly aggressive; the higher incidence of antisocial behavior was observed only for the group with both factors. As the authors of that study note, knowledge about specific genetic risk factors that make people more susceptible to particular environmental effects could strengthen multiple-risk models, such as those discussed in the previous two chapters.
97

Parental contributions to the child’s environment Obviously, a highly salient and important part of a child’s environment is the parents’ relationship with the child—the manner in which they interact with him or her, the general home environment they provide, the experiences they arrange for the child, the encouragement they offer for particular behaviors, attitudes, and activities, and so on. Less obvious is the idea that the environment that parents provide for their children is due in part to the parents’ own genetic makeup. Parents’ behavior toward their children (e.g., how warm or reserved they are, how patient or short-fused) is genetically influenced, as are the kinds of preferences, activities, and resources to which they expose their children (Plomin & Bergeman, 1991). For example, the child of a highly musical parent is likely to hear more music while growing up than are children whose parents are less musically inclined. Parents who are skilled readers and enjoy and value reading are likely to read often for pleasure and information and are likely to have lots of books around the house. They are also more likely to read frequently to their children and to take them to the library. In contrast, parents for whom reading is challenging and not a source of pleasure are less likely to provide a highly literate environment for their children (Scarr, 1992).
4. Child’s Phenotype–Child’s Environment
Relation 4 in our model restates the active child theme—the child as a source of his or her own development. As noted in Chapter 1, children are not just the passive recipients of a preexisting environment. Rather, they are active creators of the environment in which they live in two important ways. First, by virtue of their nature and behavior, they actively evoke certain kinds of responses from others (Scarr, 1992; Scarr & McCartney, 1983). Babies who enjoy being cuddled are more likely to receive cuddling than are squirmy babies. Impulsive children hear “No,” “Don’t,” “Stop,” and “Be careful” more often than inhibited children do. Indeed, the degree to which parent–child relationships are mutually responsive is largely a function of the child’s genetically influenced behavioral characteristics (Deater-Deckard & O’Connor, 2000).
The second way in which children create their own environment is by actively selecting surroundings and experiences that match their interests, talents, and personality characteristics (Scarr, 1992). As soon as infants become capable of self-locomotion, for example, they start selecting certain objects in the environment for exploration. Some very young children (especially boys) develop extremely intense interests in particular kinds of objects or activities that do not stem from parental encouragement (DeLoache, Simcock, & Macari, 2007). For example, many little boys become obsessed with vehicles and construction equipment. Other young children develop idiosyncratic and even quite peculiar interests (e.g., blenders, roadkill). For many parents, the origin of these preschool passions is totally obscure, and occasionally worrisome, because they do not realize how common these intense interests actually are.
98
Beginning in the preschool years, children’s friendship opportunities increasingly depend on their own characteristics, as they choose playmates and pals with whom they feel compatible—the “birds of a feather flock together” phenomenon. And, as noted in Chapter 1, with age, children play an ever more active role in selecting their own environment. As they gain more autonomy, they increasingly select aspects of the environment that fit their temperament and abilities. Returning to the reading example, children who enjoy reading will read more books than will children who find reading tedious. The more they read, the more skillful readers they become, leading them to choose increasingly more challenging books, which, in turn, leads them to acquire advanced vocabulary, improve their language comprehension, and enhance their general knowledge base, resulting in greater success in school.
5. Child’s Environment–Child’s Genotype
The fifth relationship in our model is perhaps the most surprising. Until fairly recently, geneticists thought of the genotype as being “fixed” at birth. But as discussed in Chapter 1, the new field of epigenetics has turned this conventional wisdom on its head. That is, it is now known that although the structure of DNA remains “fixed” (mutations aside), certain epigenetic mechanisms, mediated by the environment, can alter the functioning of genes and create stable changes in their expression—and some of these changes can be passed on to the next generation.
Epigenetic factors can help explain why identical twins do not have identical pathways through life: different environments can alter gene expression in subtle ways across developmental time. These stable changes in gene expression that are mediated by the environment involve processes of methylation, which silence gene expression. Differences in experience over the course of development are reflected in differences in methylation levels. Consider identical twin pairs at the age of 3 and at the age of 50, for example. Three-year-old co-twins have had highly overlapping life experiences, whereas many 50-year-old co-twins are likely to have had a far more divergent range of experiences. In a study that measured differences in DNA methylation levels in 3- and 50-year-old identical co-twins, researchers found that, whereas there were virtually no differences in the 3-year-olds’ levels, roughly one-third of the 50-year-olds showed “remarkable” differences—and the greater the differences in the twins’ lifestyle and experiences, the greater the differences in their methylation levels (Fraga et al., 2005).
How might the environment exert its effects through epigenetic mechanisms? To date, the bulk of the behavioral research on this topic has focused on nonhuman animal models, with clear evidence that low-quality maternal care has epigenetic effects, permanently changing the animal’s pattern of gene expression (for a recent review, see van IJzendoorn, Bakermans–Kranenburg, & Ebstein, 2011). In particular, poor maternal care affects the methylation of genes involved in glucocorticoid receptors, which influence how the animal copes with stress (e.g., T.-Y. Zhang & Meaney, 2010). As you saw in Chapter 1, there is emerging evidence suggesting similar effects of early stress on methylation in humans (e.g., Essex et al., 2013). The myriad risk factors associated with growing up in poverty appear to act on developing children via epigenetic processes as well; adults who grew up in impoverished households exhibit different patterns of gene expression decades later than do adults who grew up in high-SES homes, regardless of their SES as adults (e.g., G. E. Miller et al., 2009).
99
Our discussion of the five kinds of gene–environment interactions has emphasized the myriad challenges in understanding how genes function in the development of individuals. Nevertheless, the conceptualization we have presented is greatly simplified. This is particularly true for the fifth relationship—epigenetics—which, when considered in full, suggests that the line between genes and environment is blurry at best. The complexity of gene–environment relationships raises both challenges and opportunities for developmental scientists. One challenge is that the genome can no longer comfortably be considered immutable irrespective of the widely varying environments in which children develop. One opportunity is that as this field continues to develop, it may become possible to determine which aspects of the environment are most likely to have a lasting impact on children’s eventual health and well-being.
Behavior Genetics
behavior genetics the science concerned with how variation in behavior and development results from the combination of genetic and environmental factors
The rapidly expanding field known as behavior genetics is concerned with how variation in behavior and development results from the interaction of genetic and environmental factors. Behavior geneticists ask the same sort of question Galton asked about eminence: “Why are people different from one another?” Why, in any group of human beings, do we vary in terms of how smart, sociable, depressed, aggressive, and religious we are? The answer given by behavior geneticists is that all behavioral traits are heritable; that is, they are all influenced to some degree by hereditary factors (Bouchard, 2004; Turkheimer, 2000). As noted, the kind of traits that have been of particular interest to behavior geneticists—intelligence, sociability, mood, aggression, and the like—are polygenic, that is, affected by the combination of many genes. They are also multifactorial, that is, affected by a host of environmental factors as well as genetic ones. Thus, the potential sources of variation are vast.
multifactorial refers to traits that are affected by a host of environmental factors as well as genetic ones
To fully answer Galton’s question, behavior geneticists try to tease apart genetic and environmental contributions to the differences observed among a population of people or other animals. Two premises underlie this endeavor:
- To the extent that genetic factors are important for a given trait or behavior, individuals who are genotypically similar should be phenotypically similar. In other words, behavior patterns should “run in families”: children should be more similar to their parents and siblings than to second- or third-degree relatives or unrelated individuals.
heritable refers to any characteristics or traits that are influenced by heredity
- To the extent that shared environmental factors are important, individuals who were reared together should be more similar than people who were reared apart.
Behavior Genetic Research Designs
As it was for Galton, the mainstay of modern behavior-genetics research is the family study. In order to examine genetic and environmental contributions to a given trait or characteristic, behavior geneticists first measure that trait in people who vary in terms of genetic relatedness—parents and their children, identical and fraternal twins, nontwin siblings, and so on. Next, they assess how highly correlated the measures of the trait are among individuals who vary in the degree to which they are genetically related. (As you may recall from Chapter 1, the strength and direction of a correlation express the extent to which two variables are related; the higher the correlation, the more precisely scores on one variable can be predicted from scores on the other.) Finally, behavior geneticists compare the resulting correlations to see if they are (1) higher for more closely related individuals than for less closely related people, and (2) higher for individuals who share the same environment than for individuals who do not.
100

There are several specialized family-study designs that are particularly helpful in assessing genetic and environmental influences. One is the twin-study design, which compares the correlations for identical (monozygotic, or MZ) twins with those for same-sex fraternal (dizygotic, or DZ) twins. As you will recall, identical twins have 100% of their genes in common (though the expression of these genes are affected by epigenetic factors over the course of development, as discussed in the previous section), whereas fraternal twins are only 50% genetically similar (just like nontwin siblings). For twins who grow up together, the degree of similarity of the environment is generally assumed to be equal. Both types of twins shared the same womb, were born at the same time, have lived in the same family and community, and are always the same age when tested. Thus, with different levels of genetic similarity and essentially equal environmental similarity, the difference between the correlations for the two types of twins is treated as an index of the importance of genetic factors. If the correlation between identical twins on a given trait or behavior is substantially higher than that between fraternal twins, it is assumed that genetic factors are substantially responsible for the difference.
Another family-study design used for assessing genetic and environmental influences is the adoption study. In this approach, researchers examine whether adopted children’s scores on a given measure are correlated more highly with those of their biological parents and siblings or with those of their adoptive parents and siblings. Genetic influences are inferred to the extent that children resemble their biological relatives more than they do their adoptive ones.
The ideal behavior-genetics design—the adoptive twin study—compares identical twins who grew up together versus identical twins who were separated shortly after birth and raised apart. If the correlations for twins reared apart are similar to those for twins reared together, it suggests that environmental factors have little effect. Conversely, to the extent that the correlations between identical twins who grew up in different environments are lower than those for identical twins who grew up together, environmental influence is inferred. Box 3.2 describes some of the remarkable findings that have emerged from studies of twins reared apart, as well as some of the problems with such research.
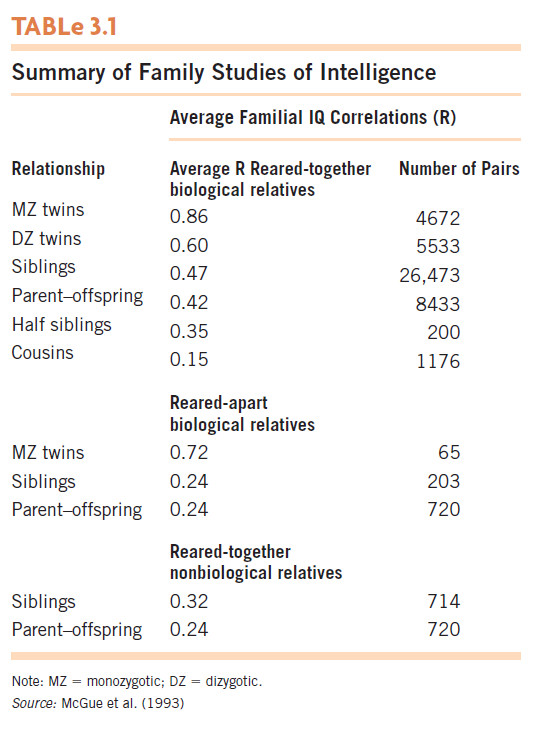
Family studies of intelligence The most common focus of behavior-genetics family studies has been intelligence. Table 3.1 summarizes the results of more than 100 family studies of IQ through adolescence. The pattern of results reveals both genetic and environmental influences. Genetic influence is shown by generally higher correlations for higher degrees of genetic similarity. Most notable is the finding that identical (MZ) twins resemble one another in IQ more than do same-sex fraternal (DZ) twins. At the same time, environmental influences are reflected in the fact that iden-tical twins are not identical in terms of IQ. Further evidence for an environmental role is that MZ twins who are reared together are more similar than those reared apart.
101
Box 3.2: individual differences: Identical Twins Reared Apart
Video: Identical Twins: Growing Up Apart

Oskar Stohr and Jack Yufa are identical twins who were separated shortly after their birth in Trinidad. Oskar was raised by his grandmother in Germany as a Catholic and a Nazi. Jack was raised by his father, in the Caribbean, as a Jew. Despite their very different backgrounds, when the brothers first met as middle-aged men recruited for a research study in Minneapolis, they discovered a remarkable number of similarities between them:
They like spicy foods and sweet liqueurs, are absent-minded, have a habit of falling asleep in front of the television, think it’s funny to sneeze in a crowd of strangers, flush the toilet before using it, store rubber bands on their wrists, read magazines back to front, dip buttered toast in their coffee. Oskar is domineering toward women and yells at his wife, which Jack did before he was separated.
(Holden, 1980, p. 1324)

Jack and Oskar are participants in the Minnesota Study of Twins Reared Apart, an extensive research project on identical twins separated early in life (Bouchard et al., 1990). More than 100 pairs of such twins have been located, recruited for the study, and brought to Minneapolis to undergo an extensive battery of physiological and psychological tests. Many twin siblings were meeting for the first time since infancy. (The reunited twins in the photo at right showed almost as many striking similarities as did Jack and Oskar, including their having held several very similar jobs and being volunteer firemen.) The motivation for this large-scale study is to examine genetic and environmental contributions to development and behavior by comparing individuals who are genetically identical but who grew up in different environments.
The Minnesota team of investigators has been struck by the extent of the similarities they have found in the separated twins; they have identified genetic contributions to “almost every behavioral trait so far investigated from reaction time to religiosity” (Bouchard et al., 1990).
As striking as the similarities between separated twins may be, there are several problems with automatically assuming that these similarities are attributable to genetic factors. One issue is that it would be a great oversimplification to suggest that all of the similar traits shared by separated twins are genetic. For example, it would be a stretch to argue that the men in the photograph share a set of genes that predetermined that they both would become firemen. As previously noted, genes code for proteins, not for anything as complex as an occupation (or choice of facial hair). An additional issue is the practice of selective placement: adoption agencies generally try to place children with families of the same general background and race, so the environments of the separated siblings are often similar in many ways. It is extremely rare for separated twins to be raised like Jack and Oskar, with different languages, religions, and cultures. In fact, the majority of the twins in most behavior-genetics studies are from predominantly White, middle-class families in Western countries. As behavior geneticists Levine and Suzuki (1993) commented:
Take one of those kids and put him in a really different environment, like in a family of bushmen in Africa, or in a farming village in mainland China, and then come back twenty years later and see if you find two firemen who dress the same!
(p. 241)
102
Does the relative influence that genes and environment have on intelligence change over the course of development? One might expect that as children get older and have ever more (and more varied) experiences in the world, genetic influences on IQ would decrease. Surprisingly, recent studies have revealed exactly the opposite pattern: as twins get older, the degree of variance in IQ accounted for by their genetic similarity increases. In a study of 11,000 MZ and DZ twin pairs across four countries, researchers found that the correlations in IQ between co-twins increased with age for MZ twins and decreased with age for DZ twins. These divergent patterns were observed first from childhood to adolescence, and again from adolescence to young adulthood (Haworth et al., 2009). The same pattern of results was revealed in a large longitudinal study that compared MZ and DZ twin pairs in early childhood (2- to 4-year-olds) and middle childhood (7- to 10-year-olds): for the younger children, shared environment accounted for more variance than did shared genes, with the opposite pattern observed for the older children (O. S. P. Davis, Haworth, & Plomin, 2009).
This surprising pattern of results—namely, that genetic influences increase with age—is consistent with the idea that people actively construct their own environment: the phenotype–environment correlation (Relation 3) discussed earlier (McGue et al., 1993; Scarr & McCartney, 1983). As children get older, they increasingly control their own experiences, and their parents have less influence over their activities. The effects of education may be particularly relevant to this pattern of results, given that educational experiences and achievements influence children’s performance on measurements of intelligence. Younger children have little or no choice about their educational setting and opportunities, whereas older children, teens, and young adults have increasingly greater choices with regard to their educational experiences (choosing more or less challenging courses of study, more or less academically oriented peer groups, and so on). It may be that identical twins’ IQs become more similar into adulthood because their common genetic predispositions lead them to select similar intellectual stimulation, whereas the IQs of fraternal twins become increasingly dissimilar because they choose divergent experiences for themselves (Scarr & McCartney, 1983).
Heritability
heritability a statistical estimate of the proportion of the measured variance on a trait among individuals in a given population that is attributable to genetic differences among those individuals
In their approach to the nature–nurture question, many behavior geneticists attempt to quantify the degree to which genes contribute to various traits. To estimate how much of the variability in measures of a given trait is attributable to genetic and environmental factors, they derive heritability estimates from correlations of the type shown in Table 3.1. Heritability is a statistical estimate of how much of the measured variance on a trait among individuals in a given population is attributable to genetic differences among those individuals.
A crucial point to understand about heritability estimates is that they tell us nothing about the relative contributions of genetic and environmental factors to the development of an individual. Instead, they estimate how much of the variation among a given population of people is due to differences in their genes. The heritability estimate for intelligence, for example, is generally considered to be approximately 50% (Bouchard, 2004; Plomin, 1990). This means that, for the population studied, roughly 50% of the variation in IQ scores is due to genetic differences among the members of the population. (It does not mean that 50% of your IQ score is due to your genetic makeup and 50% is due to your experience.) Note that this heritability estimate indicates that the environmental contribution to the variation in IQ is also approximately 50%.
103
Behavior-genetic analyses have been applied to many diverse aspects of human behavior, several of which you will encounter in other chapters of this book. To cite just a few examples, substantial heritability has been reported for infant activity level (Saudino & Eaton, 1991), temperament (Goldsmith, Buss, & Lemery, 1997), reading disability (DeFries & Gillis, 1993), and antisocial behavior (Gottesman & Goldsmith, 1994). Substantial heritability has even been reported for divorce (McGue & Lykken, 1992), TV viewing (Plomin et al., 1990), and other factors that previously seemed more likely to be influenced by the environment than by genetics (e.g., Jaffee & Price, 2007).
The implausibility of “divorcer” or “couch potato” genes brings us back to a point we made earlier: despite the common use of the phrase, there are no genes “for” particular behavior patterns. As have we stressed, genes do nothing more than code for proteins, so they affect behavior only insofar as those proteins affect the sensory, neural, and other physiological processes involved in behavior. Thus, the heritability estimate for divorce may be related to a genetic predisposition to, say, seek out novelty, and the estimate for TV viewing may be related to a genetically based low activity level or short attention span.
Heritability estimates have been criticized, both from within psychology (e.g., G. Gottlieb, Wahlsten, & Lickliter, 1998; Lerner, 1995) and from outside it (e.g., J. S. Levine & Suzuki, 1993; Lewontin, 1982). Part of the criticism stems from ways that the term “heritability” is often misinterpreted or misused by the public. One very common misuse involves the application of the concept of heritability to individuals, despite the fact that, as we have emphasized, heritability applies only to populations.
In addition, a heritability estimate applies only to a particular population living in a particular environment. Consider the case of height. Research conducted almost exclusively with North Americans and Europeans—most of them White and adequately nourished—puts the heritability of height at around 90%. But what if some segment of this population had experienced a severe famine during childhood, while the rest remained well fed? Would the heritability estimate for height still be 90%? No—because the variability due to environmental factors (poor nutrition) would increase dramatically and, therefore, the variability that could be attributed to genetic factors would decrease to the same degree. The principle of variable heritability also appeared in the discussion of IQ correlations earlier in this chapter, with the heritability estimates derived from them differing for the same individuals at different points in development (O. S. P. Davis et al., 2009).
Furthermore, it is known that heritability estimates can differ markedly for groups of people who grow up in very different economic circumstances. In the United States, for instance, heritability estimates differ considerably as a function of socioeconomic status (SES), as shown by a large twin study that included families across the SES spectrum (Turkheimer et al., 2003). In this study, almost 60% of the variance in IQ among a sample of 7-year-olds living in poverty was accounted for by shared environment, with almost none of it attributable to genetic similarity. Affluent families follow the opposite pattern, with genetic factors contributing more than environmental ones. In a related study focused on the test scores of adolescent twins, the same pattern was observed: environmental factors trumped genetic factors for poorer teens, while genetic factors trumped environmental factors for wealthier teens (Harden, Turkheimer, & Loehlin, 2007). Although it is not fully clear what causes these differing levels of heritability, both studies suggest that qualitatively different developmental forces may be operating in poor versus affluent environments.
104
A related, frequently misunderstood point is that high heritability does not imply immutability. The fact that a trait is highly heritable does not mean that there is little point in trying to improve the course of development related to that trait. Thus, for example, the fact that the heritability estimate for IQ is relatively high does not mean that the intellectual performance of young children living in poverty cannot be improved by appropriate intervention efforts (see Chapter 8).
Finally, because they are relevant only within a given population, heritability estimates tell us nothing about differences between groups. The heritability score for IQ, for example, provides little insight into the meaning of differences in the IQ scores of different groups of Americans. European Americans, on average, score 15 points higher on IQ tests than do African Americans. Some people mistakenly assume that because IQ is estimated to be 50% heritable, the difference between these two groups’ IQ scores is genetically based. This assumption is unwarranted, given the large overall disparities between the two groups in family income and education, quality of neighborhood schools, health care, and myriad other factors.
Environmental Effects
Every examination of genetic contributions to behavior and development is also, necessarily, a study of environmental influences: estimating heritability automatically estimates the proportion of variance not attributable to genes. Because heritability estimates rarely exceed 50%, a large contribution from environmental factors is usually indicated.
Behavior geneticists try to assess the extent to which aspects of an environment shared by biologically related people make them more alike and to what extent nonshared experiences make them different. The most obvious source of shared environment is growing up in the same family. Shared-environment effects can also be inferred when twins or other relatives are more similar on some trait than would be expected on the basis of their genetic relatedness. For example, substantial shared-environmental influence has been inferred for positive emotion in toddlers and young children because fraternal and identical twins who were reared together were equally similar in the degree to which they showed pleasure (Goldsmith et al., 1997). Shared-environment effects are also being discovered for disorders that have a clear genetic component. For instance, as discussed in Box 3.1, twin studies of autism spectrum disorder (ASD) have consistently provided evidence for genetic effects (with heritability of the disorder being greater for MZ twins than for DZ twins). However, in a recent large-scale study of twin pairs in which at least one co-twin had an ASD diagnosis, researchers found a substantial shared-environment effect on the likelihood that the second twin also had an ASD diagnosis (Hallmayer et al., 2011).

105
Surprisingly, behavior geneticists have reported little evidence of shared-environment effects for some other aspects of development. For example, with respect to personality, the correlations for adoptive siblings are often near zero (D. C. Rowe, 1994). The same is true for some types of psychopathology, including schizophrenia (Gottesman, 1991). As noted in Chapter 1, being adopted into a family with a schizophrenic sibling does not increase the risk that a child will become schizophrenic. In addition, the risk of schizophrenia is the same for the biological child of a schizophrenic parent regardless of whether the child is raised by that parent or is adopted away at birth (Kety et al., 1994).
Behavior geneticists’ investigations of the effects of nonshared environments arise from the recognition that even children who grow up in the same family do not have all their experiences in common—either inside or outside the family. Within the family, birth order may result in quite different experiences for siblings. The oldest child in a large family, for example, may have been reared by young, energetic, but inexperienced, parents, whereas that child’s much younger sibling will be parented by older and more sedentary, but more knowledgeable, individuals who are likely to have more resources available than they did as first-time parents. In addition, as discussed in Chapter 1, siblings may experience their parents’ behavior toward them differently (the “Mom always loved you best” syndrome). They may also be affected quite differently by an event they experience in common, such as the divorce of their parents (Hetherington & Clingempeel, 1992). Finally, siblings may be highly motivated to differentiate themselves from one another (Sulloway, 1996). The younger sibling of a star student may strive to be a star athlete instead, and a child who observes a sibling disappearing into a self-destructive pattern of drug and alcohol abuse may become determined to follow a different path. As these examples illustrate, siblings themselves are an important part of the environment, and each provides a different constellation of experiences for the others. This is another factor that makes each child’s experience within the family different.
Outside the family, siblings can also have highly divergent experiences, partly as a result of belonging to different peer groups. Highly active siblings who both like physical challenges and thrills will have very different experiences if one takes up rock climbing while the other hangs out with delinquents. Idiosyncratic life events—suffering a serious accident, having an inspiring teacher, being bullied on the playground—can contribute further to making siblings develop differently. The primary effect of nonshared environmental factors is to increase the differences among family members (Plomin & Daniels, 1987).
review:
The five relations shown in Figure 3.1 depict the complex interplay of genetic and environmental forces in development. (1) The course of children’s development is influenced by the genetic heritage they receive from their mother and father, with their sex determined solely by their father’s chromosomal contribution. (2) The relation between children’s genotype and phenotype depends in part on dominance patterns in the expression of some genes, but most traits of primary interest to behavioral scientists are influenced by multiple genes (polygenic inheritance). (3) As the concept of norm of reaction specifies, any given genotype will develop differently in different environments. A particularly salient part of children’s environment is their parents, including their parents’ own genetic makeup, which influences how parents behave toward their children. (4) Children’s own genetic makeup influences how they select and shape their own environment and the experiences they have in it. (5) Conversely, children’s experiences can change their genetic expression through epigenetic mechanisms.
106
The field of behavior genetics is concerned with how development results from the interaction of genetic and environmental factors. Using the family-study methodology, behavior geneticists compare the correlations among individuals who vary in the degree of genetic relatedness and in similarity of their rearing environments. Heritability estimates indicate the proportion of the variance among individuals in a given population on a given trait that is attributable to genetic differences among them. Most behavioral traits that have been measured show substantial heritability; at the same time, heritability estimates reveal the close partnership of heredity and environment in development and the fallacy of considering the influences of nature and nurture as independent of one another.