40.2 Peptidyl Transferase Catalyzes Peptide-Bond Synthesis
✓ 5 Explain the role of the ribosome in protein synthesis.
With both the P site and the A site occupied by aminoacyl-tRNA, the stage is set for the formation of a peptide bond: the formylmethionine molecule linked to the initiator tRNA will be transferred to the amino group of the amino acid in the A site. The formation of the peptide bond, one of the most important reactions in life, is a thermodynamically spontaneous reaction catalyzed by a site on the 23S rRNA of the 50S subunit called the peptidyl transferase center. This catalytic center is located deep in the 50S subunit near the tunnel that allows the nascent peptide to leave the ribosome.
The ribosome, which enhances the rate of peptide bond synthesis by a factor of 107 over the uncatalyzed reaction (∼10−4 M−1 s−1), derives much of its catalytic power from catalysis by proximity and orientation. The ribosome positions and orients the two substrates so that they are situated to take advantage of the inherent reactivity of an amine group (on the aminoacyl-tRNA in the A site) with an ester (on the initiator tRNA in the P site).
The reaction begins when the amino group of the aminoacyl-tRNA in the A site, in its unprotonated state, makes a nucleophilic attack on the ester linkage between the initiator tRNA and the formylmethionine molecule in the P site (Figure 40.7A). The nature of the transition state that follows the attack is not established, and several models are plausible. One model proposes roles for the 2′OH of the adenosine of the tRNA in the P site and a molecule of water at the peptidyl transferase center (Figure 40.7B). The nucleophilic attack of the α-amino group generates an eight-membered transition state in which three protons are shuttled about in a concerted manner. The proton of the attacking amino group hydrogen bonds to the 2′ oxygen of ribose of the tRNA. The hydrogen of 2′OH in turn interacts with the oxygen of the water molecule at the center, which then donates a proton to the carbonyl oxygen. Collapse of the transition state with the formation of the peptide bond allows protonation of the 3′OH of the now-empty tRNA in the P site (Figure 40.7C). The stage is now set for translocation and formation of the next peptide bond.
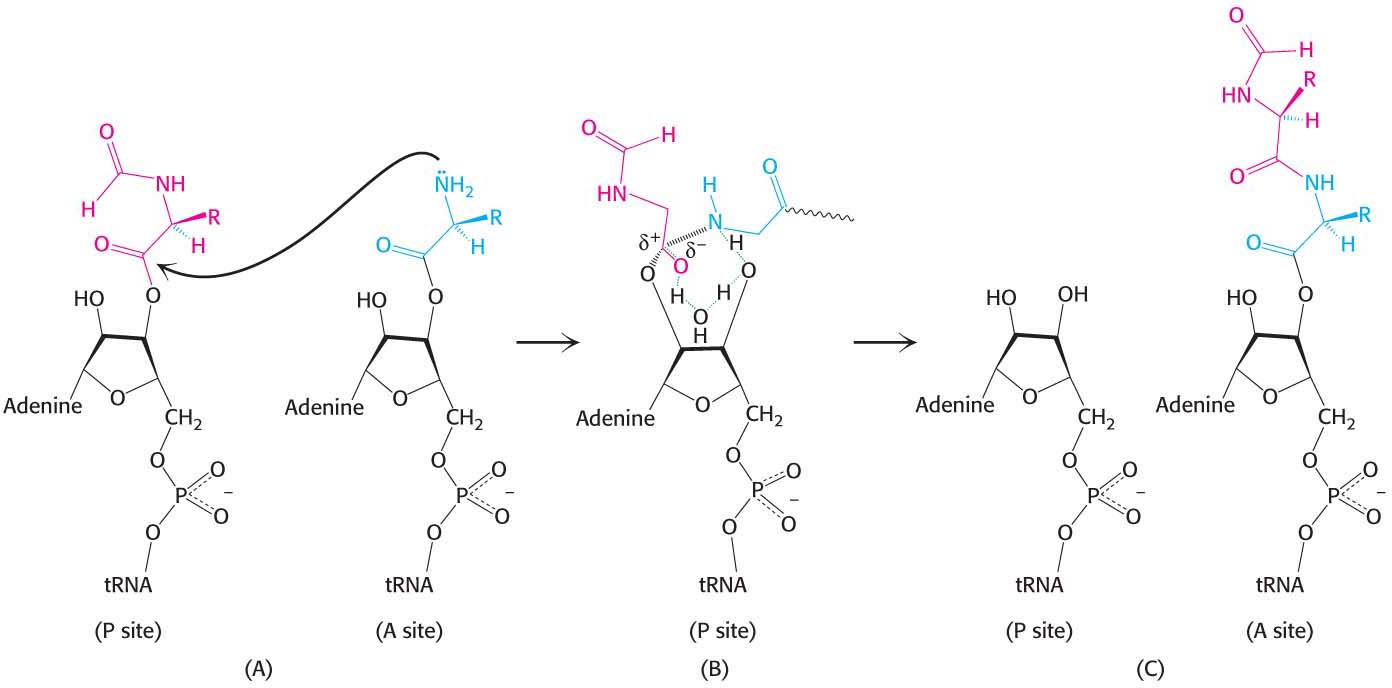
Figure 40.7: Peptide-bond formation. (A) The amino group of the aminoacyl-tRNA attacks the carbonyl group of the ester linkage of the peptidyl-tRNA. (B) An eight-membered transition state is formed. Note: Not all atoms are shown, and some bond lengths are exaggerated for clarity. (C) This transition state collapses to form the peptide bond and release the deacylated tRNA.
The Formation of a Peptide Bond Is Followed by the GTP-Driven Translocation of tRNAs and mRNA
With the formation of the peptide bond, the peptide chain is now attached to the tRNA whose anticodon is in the A site on the 30S subunit. The two ribosomal subunits rotate with respect to one another, and this structural change places the CCA end of the same tRNA and its peptide in the P site of the large subunit (Figure 40.8). However, protein synthesis cannot continue without the translocation of the mRNA and the tRNAs within the ribosome. The mRNA must move by a distance of three nucleotides so that the next codon is positioned in the A site for interaction with the incoming aminoacyl-tRNA. The peptidyl-tRNA moves out of the A site into the P site on the 30S subunit and at the same time, the deacylated tRNA moves out of the P site into the E site and is subsequently released from the ribosome. The movement of the peptidyl-tRNA into the P site shifts the mRNA by one codon, exposing the next codon to be translated in the A site.
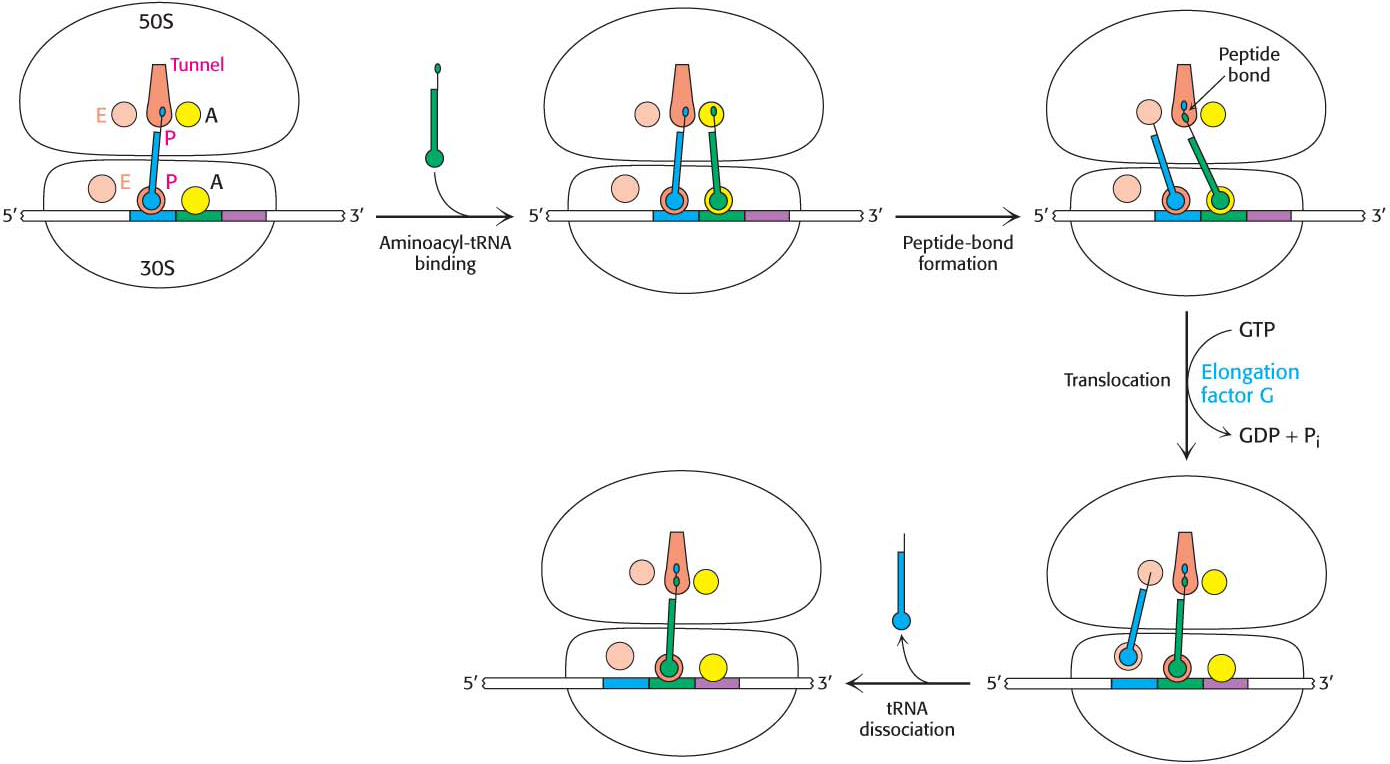
Figure 40.8: Mechanism of protein synthesis. The cycle begins with peptidyl-tRNA in the P site. An aminoacyl-tRNA binds in the A site. With both sites occupied, a new peptide bond is formed. The tRNAs and the mRNA are translocated through the action of elongation factor G, which moves the deacylated tRNA to the E site. When there, the tRNA is free to dissociate to complete the cycle.
The three-dimensional structure of the ribosome undergoes significant change during translocation, and evidence suggests that translocation may result from properties of the ribosome itself. However, protein factors accelerate the process. Translocation is enhanced by elongation factor G (EF-G, also called translocase). A possible mechanism for accelerating the translocation process is shown in Figure 40.9. First, EF-G in the GTP form binds to the ribosome near the A site, interacting with the 23S rRNA of the 50S subunit. The binding of EF-G to the ribosome stimulates the GTPase activity of EF-G. On GTP hydrolysis, EF-G undergoes a conformational change that displaces the peptidyl-tRNA in the A site to the P site, which carries the mRNA and the deacylated tRNA with it. The dissociation of EF-G leaves the ribosome ready to accept the next aminoacyl-tRNA into the A site.
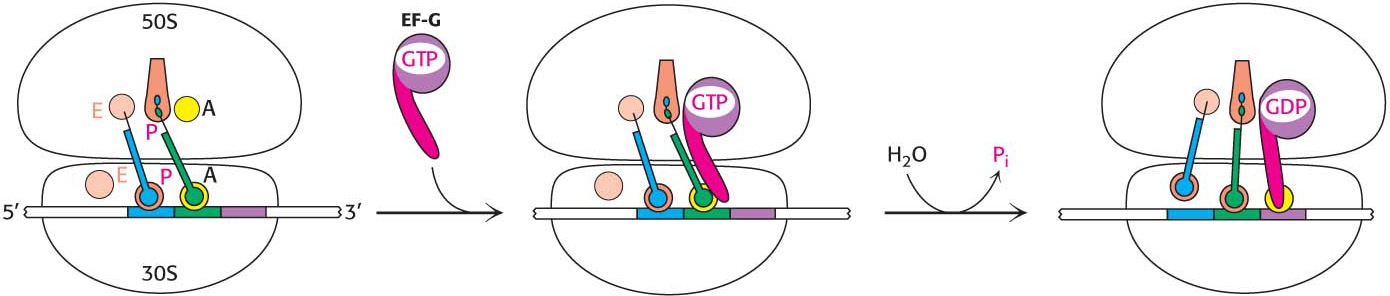
Figure 40.9: Translocation mechanism. In the GTP form, EF-G binds to the EF-Tu-binding site on the 50S subunit. This binding stimulates GTP hydrolysis, inducing a conformational change in EF-G that forces the tRNAs and the mRNA to move through the ribosome by a distance corresponding to one codon.
Note that the peptide chain remains in the P site on the 50S subunit throughout this cycle, growing into the exit tunnel. This cycle is repeated as new aminoacyl-tRNAs move into the A site, allowing the polypeptide to be elongated indefinitely. Keep in mind that all of this activity is taking place 40 times per second. Note that the polypeptide is synthesized in the amino-terminal-to-carboxyl-terminal direction (Figure 40.10). Many ribosomes can be translating an mRNA molecule simultaneously, forming a polyribosome, or a polysome (Figure 39.9).
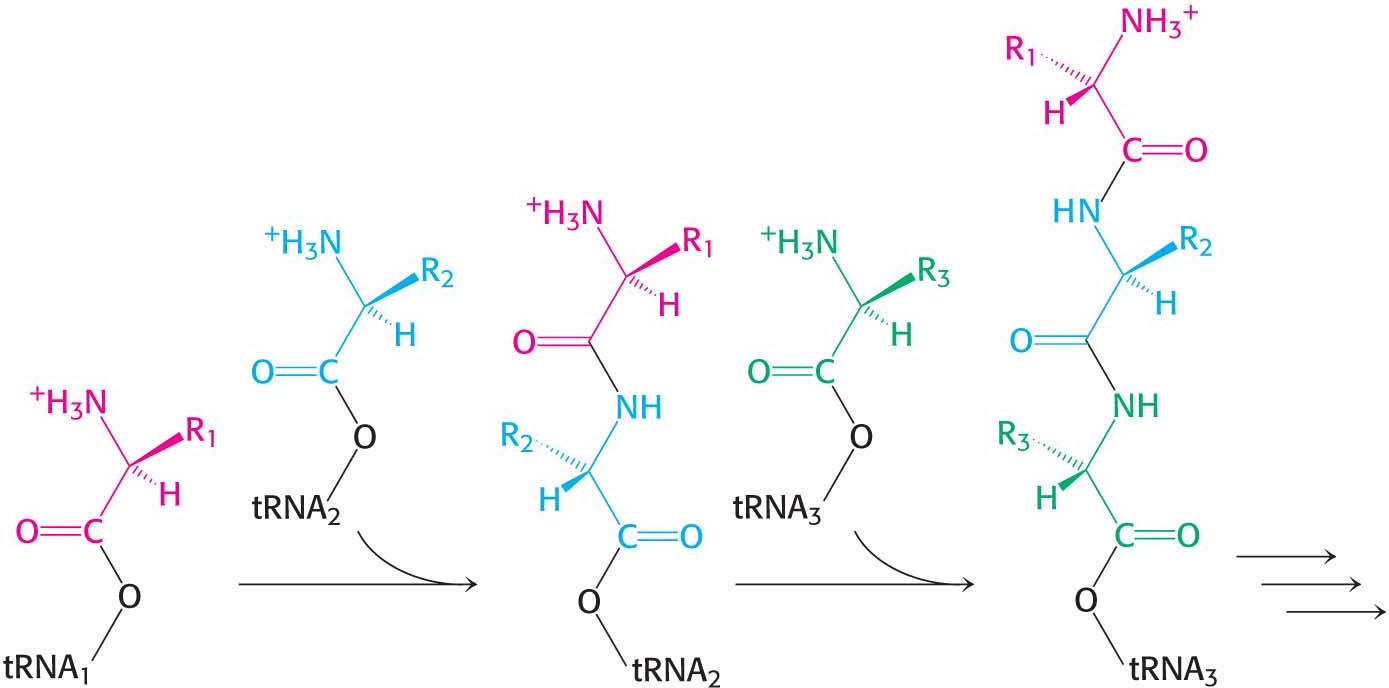
Figure 40.10: Polypeptide-chain growth. Proteins are synthesized by the successive addition of amino acids to the carboxyl terminus.
Protein Synthesis Is Terminated by Release Factors That Read Stop Codons
The final phase of translation is termination. How does the synthesis of a polypeptide chain come to an end when a Stop codon is encountered? No tRNAs with anticodons complementary to the Stop codons—UAA, UGA, or UAG—exist in normal cells. Instead, these Stop codons are recognized by proteins called release factors (RFs). One of these release factors, RF1, recognizes UAA or UAG. A second factor, RF2, recognizes UAA or UGA. A third factor, RF3, another GTPase, catalyzes the removal of RF1 or RF2 from the ribosome upon release of the newly synthesized protein.
!quickquiz! QUICK QUIZ 1
What are the roles of the protein factors required for protein synthesis?
RF1 and RF2 are compact proteins that, when bound to the ribosome, unfold to bridge the gap between the Stop codon on the mRNA and the peptidyl transferase center on the 50S subunit (Figure 40.11). The RF interacts with the peptidyl transferase center with a loop containing a highly conserved glycine-glycine-glutamine (GGQ) sequence, with the glutamine methylated on the amide nitrogen atom of the R group. This modified glutamine is crucial in promoting, assisted by the peptidyl transferase, a water molecule’s attack on the ester linkage between the tRNA and the polypeptide chain, freeing the polypeptide chain. The detached polypeptide leaves the ribosome. Transfer RNA and messenger RNA remain briefly attached to the 70S ribosome until the entire complex is dissociated through the hydrolysis of GTP in response to the binding of EF-G and another factor, called the ribosome release factor (RRF).
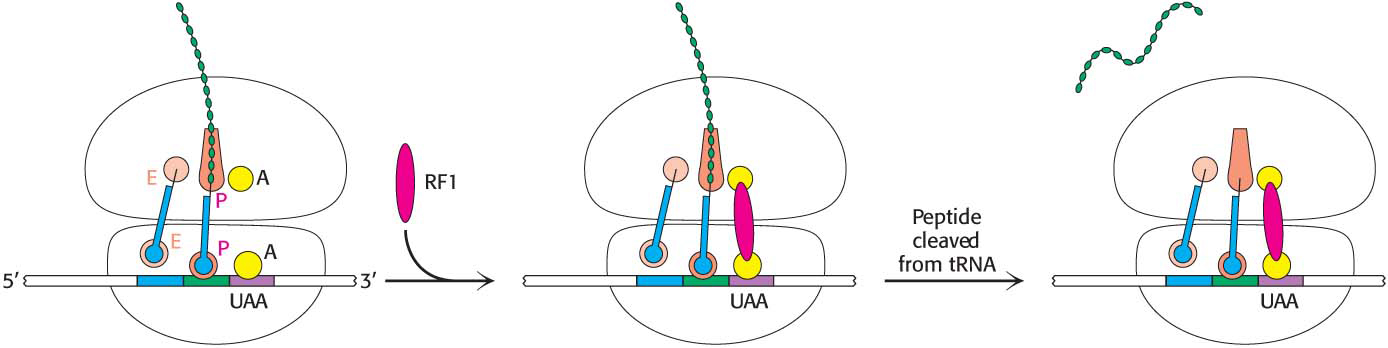
Figure 40.11: Termination of protein synthesis. A release factor recognizes a Stop codon in the A site and stimulates the release of the completed protein from the tRNA in the P site.