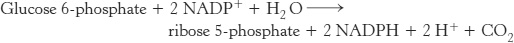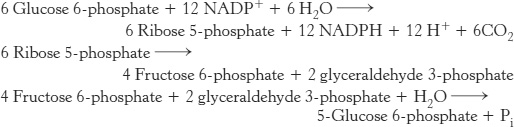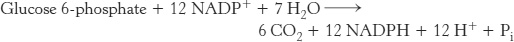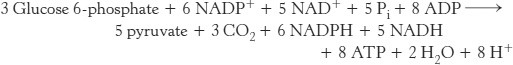The flow of glucose 6-phosphate depends on the need for NADPH, ribose 5-phosphate, and ATP
We can grasp the intricate interplay between glycolysis and the pentose phosphate pathway by examining the metabolism of glucose 6-phosphate in four different metabolic situations (Figure 20.23).
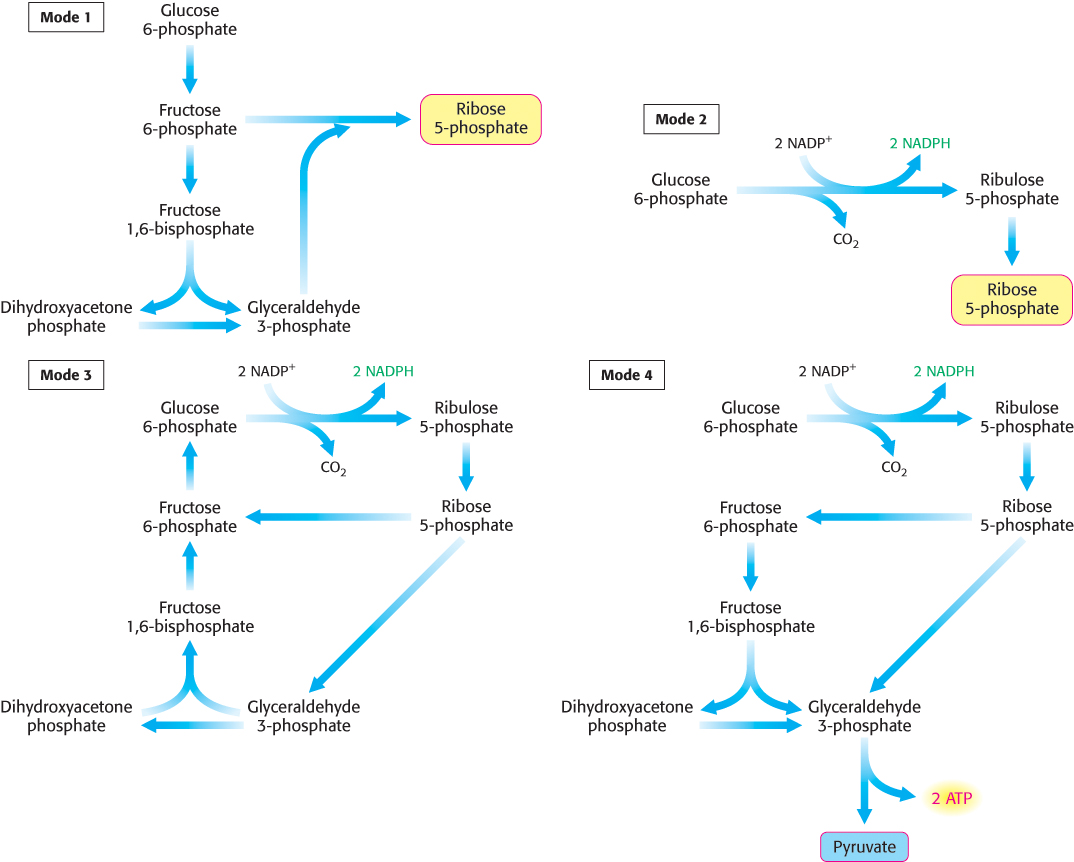
FIGURE 20.23Four modes of the pentose phosphate pathway. Major products are shown in color.
Mode 1. Much more ribose 5-phosphate than NADPH is required. For example, rapidly dividing cells need ribose 5-phosphate for the synthesis of nucleotide precursors of DNA. Most of the glucose 6-phosphate is converted into fructose 6-phosphate and glyceraldehyde 3-phosphate by the glycolytic pathway. Transaldolase and transketolase then convert two molecules of fructose 6-phosphate and one molecule of glyceraldehyde 3-phosphate into three molecules of ribose 5-phosphate by a reversal of the reactions described earlier. The stoichiometry of mode 1 is
5 Glucose 6-phosphate + ATP → 6 ribose 5-phosphate + ADP + 2 H+
Mode 2. The needs for NADPH and for ribose 5-phosphate are balanced. Under these conditions, glucose 6-phosphate is processed to one molecule of ribulose 5-phosphate while generating two molecules of NADPH. Ribulose 5-phosphate is then converted into ribose 5-phosphate. The stoichiometry of mode 2 is
Mode 3. Much more NADPH than ribose 5-phosphate is required. For example, adipose tissue requires a high level of NADPH for the synthesis of fatty acids (Table 20.4). In this case, glucose 6-phosphate is completely oxidized to CO2. Three groups of reactions are active in this situation. First, the oxidative phase of the pentose phosphate pathway forms two molecules of NADPH and one molecule of ribulose 5-phosphate. Then, ribulose 5-phosphate is converted into fructose 6-phosphate and glyceraldehyde 3-phosphate by transketolase and transaldolase. Finally, glucose 6-phosphate is resynthesized from fructose 6-phosphate and glyceraldehyde 3-phosphate by the gluconeogenic pathway. The stoichiometries of these three sets of reactions are
TABLE 20.4 Tissues with active pentose phosphate pathways
|
|
|
|
|
|
|
|
Fatty acid and cholesterol synthesis |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Maintenance of reduced glutathione |
The sum of the mode 3 reactions is
Thus, the equivalent of glucose 6-phosphate can be completely oxidized to CO2 with the concomitant generation of NADPH. In essence, ribose 5-phosphate produced by the pentose phosphate pathway is recycled into glucose 6-phosphate by transketolase, transaldolase, and some of the enzymes of the gluconeogenic pathway.
Mode 4. Both NADPH and ATP are required. Alternatively, ribulose 5-phosphate formed from glucose 6-phosphate can be converted into pyruvate. Fructose 6-phosphate and glyceraldehyde 3-phosphate derived from ribose 5-phosphate enter the glycolytic pathway rather than reverting to glucose 6-phosphate. In this mode, ATP and NADPH are concomitantly generated, and five of the six carbons of glucose 6-phosphate emerge in pyruvate.
Pyruvate formed by these reactions can be oxidized to generate more ATP or it can be used as a building block in a variety of biosyntheses.
The pentose phosphate pathway is required for rapid cell growth
Rapidly dividing cells, such as cancer cells, require ribose 5-phosphate for nucleic acid synthesis and NADPH for fatty acid synthesis, which in turn is required to form membrane lipids (Section 26.1). Recall that rapidly dividing cells switch to aerobic glycolysis to meet their ATP needs (Section 16.2). Glucose 6-phosphate and glycolytic intermediates are then used to generate NADPH and ribose 5-phosphate using the pentose phosphate pathway as described in Modes 1 and 3. The diversion of glycolytic intermediates into the nonoxidative phase is facilitated by the expression of the gene for a pyruvate kinase isozyme, PKM. PKM has a low catalytic activity, creating a bottle-neck in the glycolytic pathway. Glycolytic intermediates accumulate and are then used by the pentose phosphate pathway to synthesize NADPH and ribose 5-phosphate. The shunting of phosphorylated intermediates into the nonoxidative phase of the pentose phosphate pathway is further enabled by the inhibition of triose phosphate isomerase by phosphoenolpyruvate, the substrate of PKM.
Through the looking-glass: The Calvin cycle and the pentose phosphate pathway are mirror images
 The complexities of the Calvin cycle and the pentose phosphate pathway are easier to comprehend if we consider them as functional mirror images of each other. The Calvin cycle begins with the fixation of CO2 and proceeds to use NADPH in the synthesis of glucose. The pentose phosphate pathway begins with the oxidation of a glucose-derived carbon atom to CO2 and concomitantly generates NADPH. The regeneration phase of the Calvin cycle converts C6 and C3 molecules back into the starting material—the C5 molecule ribulose 1,5-bisphosphate. The pentose phosphate pathway converts a C5 molecule, ribulose 5-phosphate, into C6 and C3 intermediates of the glycolytic pathway. Not surprisingly, in photosynthetic organisms, many enzymes are common to the two pathways. We see the economy of evolution: the use of identical enzymes for similar reactions with different ends.
The complexities of the Calvin cycle and the pentose phosphate pathway are easier to comprehend if we consider them as functional mirror images of each other. The Calvin cycle begins with the fixation of CO2 and proceeds to use NADPH in the synthesis of glucose. The pentose phosphate pathway begins with the oxidation of a glucose-derived carbon atom to CO2 and concomitantly generates NADPH. The regeneration phase of the Calvin cycle converts C6 and C3 molecules back into the starting material—the C5 molecule ribulose 1,5-bisphosphate. The pentose phosphate pathway converts a C5 molecule, ribulose 5-phosphate, into C6 and C3 intermediates of the glycolytic pathway. Not surprisingly, in photosynthetic organisms, many enzymes are common to the two pathways. We see the economy of evolution: the use of identical enzymes for similar reactions with different ends.