35.3
Kinesin and Dynein Move Along Microtubules
In addition to actin, the cytoskeleton includes other components, notably intermediate filaments and microtubules. Microtubules serve as tracks for two classes of motor proteins—namely, kinesins and dyneins. Kinesins moving along microtubules usually carry cargo such as organelles and vesicles from the center of a cell to its periphery. Dyneins are important in sliding microtubules relative to one another during the beating of cilia and flagella on the surfaces of some eukaryotic cells. Additionally, dynein carries cargo from the cell periphery to the cell center.
 Some members of the kinesin family are crucial to the transport of organelles and other cargo to nerve endings at the peripheries of neurons. It is not surprising, then, that mutations in these kinesins can lead to nervous system disorders. For example, mutations in a kinesin called KIF1β can lead to the most common peripheral neuropathy (weakness and pain in the hands and feet), Charcot-Marie-Tooth disease, which affects 1 in 2500 people. A glutamine-to-leucine mutation in the P-loop of the motor domain of this kinesin has been found in some affected persons. Knockout mice with a disruption of the orthologous gene have been generated. Mice heterozygous for the disruption show symptoms similar to those observed in human beings; homozygotes die shortly after birth. Mutations in other kinesin genes have been linked to human spastic paraplegia. In these disorders, defects in kinesin-linked transport may impair nerve function directly, and the decrease in the activity of specific neurons may lead to other degenerative processes.
Some members of the kinesin family are crucial to the transport of organelles and other cargo to nerve endings at the peripheries of neurons. It is not surprising, then, that mutations in these kinesins can lead to nervous system disorders. For example, mutations in a kinesin called KIF1β can lead to the most common peripheral neuropathy (weakness and pain in the hands and feet), Charcot-Marie-Tooth disease, which affects 1 in 2500 people. A glutamine-to-leucine mutation in the P-loop of the motor domain of this kinesin has been found in some affected persons. Knockout mice with a disruption of the orthologous gene have been generated. Mice heterozygous for the disruption show symptoms similar to those observed in human beings; homozygotes die shortly after birth. Mutations in other kinesin genes have been linked to human spastic paraplegia. In these disorders, defects in kinesin-linked transport may impair nerve function directly, and the decrease in the activity of specific neurons may lead to other degenerative processes.
Microtubules are hollow cylindrical polymers
Microtubules are a major component of the cytoskeleton. They are built from two kinds of homologous 50-kDa subunits, α- and β-tubulin, which assemble in a helical array of alternating tubulin types to form the wall of a hollow cylinder (Figure 35.20). Alternatively, a microtubule can be regarded as 13 protofilaments that run parallel to its long axis. The outer diameter of a microtubule is 30 nm, much larger than that of actin (8 nm). Like actin, microtubules are polar structures. The minus end of a microtubule is anchored near the center of a cell, whereas the plus end extends toward the cell surface.
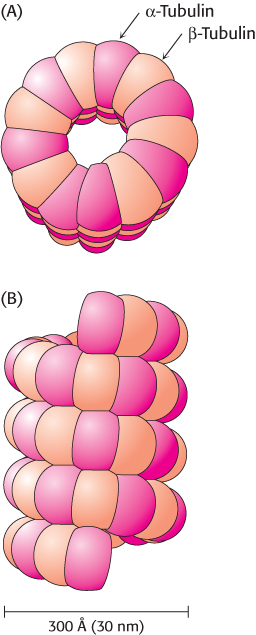
FIGURE 35.20Microtubule structure. Schematic views of the helical structure of a microtubule. α-Tubulin is shown in dark red and β-tubulin in light red. (A) Top view. (B) Side view.
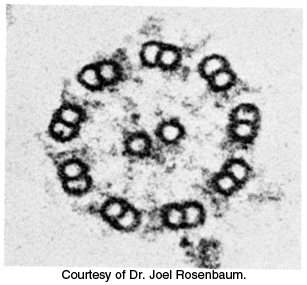
FIGURE 35.21Microtubule arrangement. Electron micrograph of a cross section of a flagellar axoneme shows nine microtubule doublets surrounding two singlets.
Microtubules are also key components of cilia and flagella present on some eukaryotic cells. For example, sperm propel themselves through the motion of flagella containing microtubules. The microtubules present in these structures adopt a common architecture (Figure 35.21). A bundle of microtubules called an axoneme is surrounded by a membrane contiguous with the plasma membrane. The axoneme is composed of a peripheral group of nine microtubule pairs surrounding two singlet microtubules. This recurring motif is often called a 9 + 2 array. Dynein drives the motion of one member of each outer pair relative to the other, causing the overall structure to bend.
Microtubules are important in determining the shapes of cells and in separating daughter chromosomes in mitosis. They are highly dynamic structures that grow through the addition of α- and β-tubulin to the ends of existing structures. Like actin, tubulins bind and hydrolyze nucleoside triphosphates, although, for tubulin, the nucleotide is GTP rather than ATP. The critical concentration for the polymerization of the GTP forms of tubulin is lower than that for the GDP forms. Thus, a newly formed microtubule consists primarily of GTP-tubulins. Through time, the GTP is hydrolyzed to GDP. The GDP-tubulin subunits in the interior length of a microtubule remain stably polymerized, whereas GDP subunits exposed at an end have a strong tendency to dissociate. Marc Kirschner and Tim Mitchison found that some microtubules in a population lengthen while others simultaneously shorten. This property, called dynamic instability, arises from random fluctuations in the number of GTP- or GDP-tubulin subunits at the plus end of the polymer. The dynamic character of microtubules is crucial for processes such as mitosis, which require the assembly and disassembly of elaborate microtubule-based structures such as spindle fibers.
 The structure of a tubulin dimer was determined at high resolution by electron crystallographic methods (Figure 35.22). As expected from their 40% sequence identity, α- and β-tubulin have very similar three- dimensional structures. Further analysis revealed that the tubulins are members of the P-loop NTPase family and contain a nucleotide-binding site adjacent to the P-loop. Tubulins are present only in eukaryotes, although a prokaryotic homolog has been found. Sequence analysis identified a prokaryotic protein called FtsZ (for filamentous temperature-sensitive mutant Z) that is quite similar to the tubulins. The homology was confirmed when the structure was determined by x-ray crystallography. Interestingly, this protein participates in bacterial cell division, forming ring-shaped structures at the constriction that arises when a cell divides. These observations suggest that tubulins may have evolved from an ancient cell-division protein.
The structure of a tubulin dimer was determined at high resolution by electron crystallographic methods (Figure 35.22). As expected from their 40% sequence identity, α- and β-tubulin have very similar three- dimensional structures. Further analysis revealed that the tubulins are members of the P-loop NTPase family and contain a nucleotide-binding site adjacent to the P-loop. Tubulins are present only in eukaryotes, although a prokaryotic homolog has been found. Sequence analysis identified a prokaryotic protein called FtsZ (for filamentous temperature-sensitive mutant Z) that is quite similar to the tubulins. The homology was confirmed when the structure was determined by x-ray crystallography. Interestingly, this protein participates in bacterial cell division, forming ring-shaped structures at the constriction that arises when a cell divides. These observations suggest that tubulins may have evolved from an ancient cell-division protein.
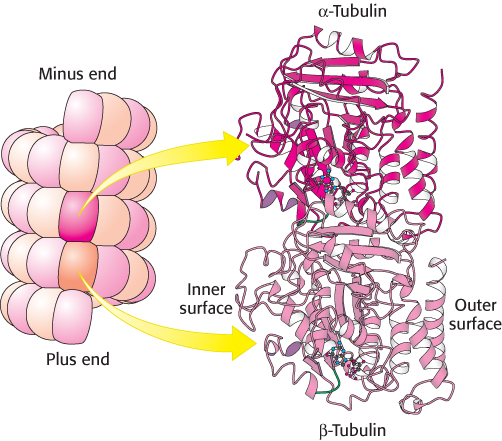
 FIGURE 35.22 Tubulin. Microtubules can be viewed as an assembly of α-tubulin–β-tubulin dimers. The structures of α-tubulin and β-tubulin are quite similar. Notice that each includes a P-loop NTPase domain (shown in purple) and a bound guanine nucleotide.
FIGURE 35.22 Tubulin. Microtubules can be viewed as an assembly of α-tubulin–β-tubulin dimers. The structures of α-tubulin and β-tubulin are quite similar. Notice that each includes a P-loop NTPase domain (shown in purple) and a bound guanine nucleotide.
[Drawn from 1JFF.pdb.]
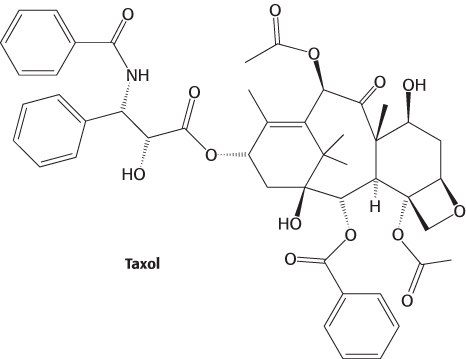
The continual lengthening and shortening of microtubules is essential to their role in cell division. Taxol, a compound isolated from the bark of the Pacific yew tree, was discovered through its ability to interfere with cell proliferation.
Taxol binds to microtubules and stabilizes the polymerized form. Taxol and its derivatives have been developed as anticancer agents because they preferentially affect rapidly dividing cells, such as those in tumors.
Kinesin motion is highly processive
Kinesins are motor proteins that move along microtubules. We have seen that muscle myosin moves along actin filaments by a process in which actin is released in each cycle; a myosin head group acting independently dissociates from actin after every power stroke. In contrast, when a kinesin molecule moves along a microtubule, the two head groups of the kinesin molecule operate in tandem: one binds, and then the next one does. A kinesin molecule may take many steps before both head groups are dissociated at the same time. In other words, the motion of kinesin is highly processive. Single-molecule measurements allow processive motion to be observed (Figure 35.23). A single kinesin molecule will typically take 100 or more steps toward the plus end of a microtubule in a period of seconds before the molecule becomes detached from the microtubule. These measurements also revealed that the average step size is approximately 80 Å, a value that corresponds to the distance between consecutive α- or β-tubulin subunits along each protofilament.
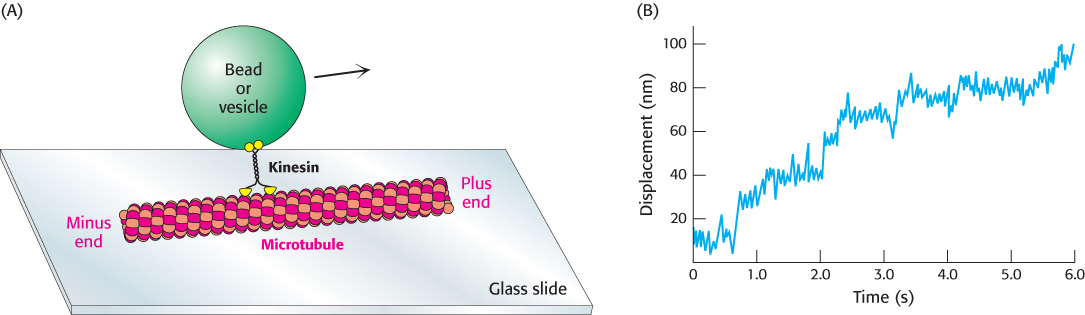
FIGURE 35.23Monitoring movements mediated by kinesin. (A) The movement of beads or vesicles, carried by individual kinesin dimers along a microtubule, can be directly observed. (B) A trace shows the displacement of a bead carried by a kinesin molecule. Multiple steps are taken in the 6-s interval. The average step size is about 8 nm (80 Å).
[(B) Data from K. Svoboda et al., Nature 365:721–727, 1993.]
An additional fact is crucial to the development of a mechanism for kinesin motion—namely, that the addition of ATP strongly increases the affinity of kinesin for microtubules. This behavior stands in contrast with the behavior of myosin; ATP binding to myosin promotes its dissociation from actin. Do these differences imply that kinesin and myosin operate by completely different mechanisms? Indeed not. Kinesin-generated movement appears to proceed by a mechanism that is quite similar to that used by myosin (Figure 35.24). Let us begin with a two-headed kinesin molecule in its ADP form, dissociated from a microtubule. Recall that the neck linker binds the head domain when ATP is bound and is released when ADP is bound. The initial interaction of one of the head domains with a tubulin dimer on a microtubule stimulates the release of ADP from this head domain and the subsequent binding of ATP. The binding of ATP triggers a conformational change in the head domain that leads to two important events. First, the affinity of the head domain for the microtubule increases, essentially locking this head domain in place. Second, the neck linker binds to the head domain. This change, transmitted through the coiled-coil domain that connects the two kinesin monomers, repositions the other head domain. In its new position, the second head domain is close to a second tubulin dimer, 80 Å along the microtubule in the direction of the plus end. Meanwhile, the intrinsic ATPase activity of the first head domain hydrolyzes the ATP to ADP and Pi. When the second head domain binds to the microtubule, the first head releases ADP and binds ATP. Again, ATP binding favors a conformational change that pulls the first domain forward. This process can continue for many cycles until, by chance, both head domains are in the ADP form simultaneously and kinesin dissociates from the microtubule. Because of the relative rates of the component reactions, a simultaneous dissociation takes place approximately every 100 cycles. Thus, kinesin “walks” hand over hand along the microtubule for approximately 100 steps before both “hands” simultaneously let go. Although recent studies have led to a structural model for the dynein power stroke, the mechanism by which it works is unknown and an interesting research frontier.

FIGURE 35.24Kinesin moving along a microtubule. (1) One head of a two-headed kinesin molecule, initially with both heads in the ADP form, binds to a microtubule. (2) The release of ADP and the binding of ATP results in a conformational change that locks the head to the microtubule and pulls the neck linker (orange) to the head domain, throwing the second domain toward the plus end of the microtubule. (3) ATP undergoes hydrolysis while the second head interacts with the microtubule. (4) The exchange of ATP for ADP in the second head pulls the first head off the microtubule, releasing Pi and moving the first domain along the microtubule. (5) The cycle repeats, moving the kinesin dimer farther down the microtubule.
Kinesin hydrolyzes ATP at a rate of approximately 80 molecules per second. Thus, given the step size of 80 Å per molecule of ATP, kinesin moves along a microtubule at a speed of 6400 Å per second. This rate is considerably lower than the maximum rate for myosin, which moves relative to actin at 80,000 Å per second. Recall, however, that myosin movement depends on the independent action of hundreds of different head domains working along the same actin filament, whereas the movement of kinesin is driven by the processive action of kinesin head groups working in pairs. Muscle myosin evolved to maximize the speed of the motion, whereas kinesin functions to achieve steady, but slower, transport in one direction along a filament.