2.2 CYTOGENETICS: CHROMOSOME MOVEMENTS DURING MITOSIS AND MEIOSIS
In 1865, Mendel presented his findings on inheritance in two lectures to the Brünn Society for the Study of the Natural Sciences, and they were then published in an obscure journal. Only about 150 copies of this journal were printed, and Mendel’s findings lay dormant for decades, to be resurrected only after his death. However, in his lifetime, Mendel was well appreciated at his monastery, was elected abbot, and managed one of the wealthiest cloisters in the land. His claim to fame was an incident in which he refused to pay a new tax imposed on the monastery by the Habsburg Empire. Mendel met the sheriff at the gate and dared him to take the keys from his pocket before he’d pay another pfennig! Of course, this is not what we know Mendel for today.
The years between 1880 and 1900 saw amazing discoveries in cytology, the study of cells, which intersected with the rediscovery of Mendel’s work. Microscopes had become more advanced, and chromosomes could be stained and visualized in the cell nucleus. Cytologists observed that, unlike other cellular components, chromosomes were meticulously divided between the two new cells during cell division. The diploid nature of somatic cells and the haploid nature of gametes were also discovered around this time. It was in this scientific environment of explosive growth in cytogenetics that, in 1900, Mendel’s principles of heredity were confirmed experimentally and rediscovered independently by three scientists: Hugo de Vries, Carl Correns, and Erich von Tschermak. The behavior of chromosomes was seen to remarkably mirror the behavior of Mendel’s hereditary particles, and the idea that the nucleus, and perhaps the chromosomes themselves, formed the basis of heredity was bandied about.
In this section, we describe the architecture of the cell and the chromosome movements that occur during somatic cell division and gamete formation, setting the stage for the chromosome theory of inheritance (the subject of Section 2.3).
Cells Contain Chromosomes and Other Internal Structures
Robert Hooke was the first to notice the cellular composition of a biological specimen, during his microscopic examination of cork in 1665 (Figure 2-7). In his famous book Micrographia, Hooke described a multitude of tiny boxes in the cork sample and coined the word cell (Latin cellula, “small compartment”). By the early 1800s, it became clear that plants are made up of cells. In 1833, Robert Brown identified the nucleus, the first subcellular structure to be discovered. In 1839, Theodor Schwann realized that animal tissue contains nuclei throughout the cells, and he proposed the cell theory, which states that all animals and plants consist of large assemblages of cells.

32
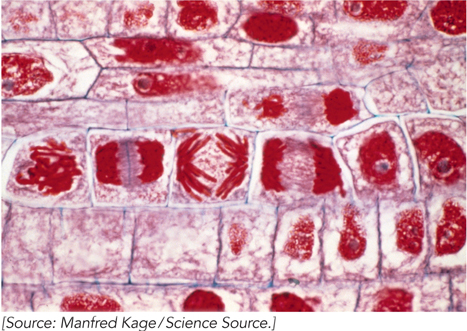
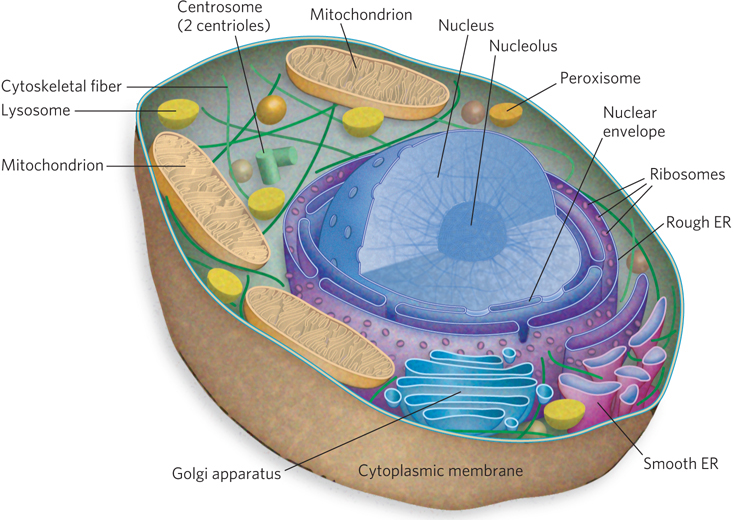
Microscopic studies of chromosomes within nuclei were first made in plant cells by Karl Wilhelm von Nägeli and Wilhelm Hofmeister between 1842 and 1849. Chromosomes were named (from the Greek for “colored body”) for their property of taking up large amounts of colored dye. The term was coined by Heinrich von Waldeyer in 1888. Figure 2-8 shows rapidly dividing cells of an onion, stained to show the chromosomes. Development of the electron microscope in 1931 eventually brought into view the detailed structure of the cell as we know it today. Each cell is bounded by the cytoplasmic membrane, encasing the cytoplasm and its variety of subcellular structures called organelles. Figure 2-9 is a schematic depiction of a typical animal cell, a eukaryotic cell; the caption describes each organelle and its function.
33
Mitosis: Cells Evenly Divide Chromosomes between New Cells
For biological information to be faithfully transmitted to daughter cells, it must be duplicated and then each complete information packet correctly partitioned into its own cell. As cells grow, they proceed through four phases, collectively known as the cell cycle: G1, S, G2, and M (Figure 2-10). In G1 phase, cells are diploid, containing two copies of each chromosome. The cellular chromosome content in G1 cells is represented as 2n, where n is the number of unique chromosomes of that species. G1 is also called the first gap, because it represents a gap in time before S phase.
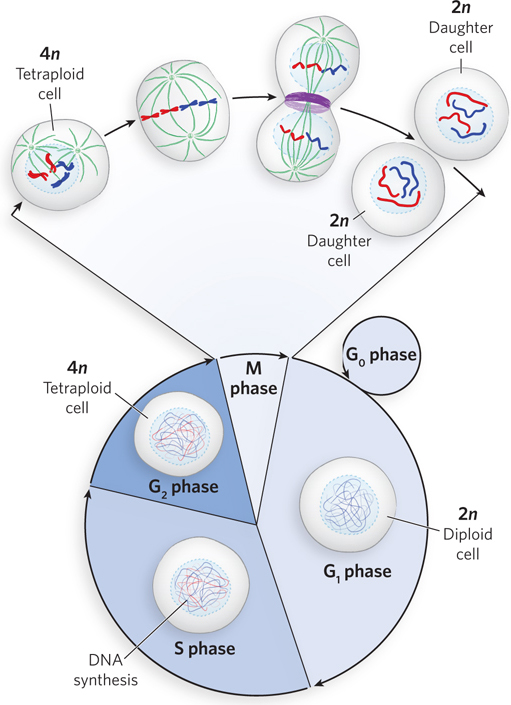
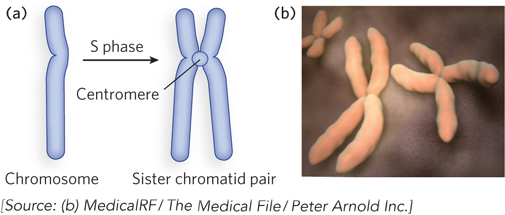
During S phase (S for synthesis), each chromosome is duplicated, and the two identical chromosomes remain together as a sister chromatid pair. The point where the sister chromatids are joined is called the centromere (Figure 2-11). At the end of S phase, each homologous chromosome exists as a sister chromatid pair, and thus the cell now contains four copies of each chromosome in the form of two sister chromatid pairs (i.e., the cell is 4n, or tetraploid). The cell next enters G2 phase, or the second gap in time, after S phase.
The final phase of the cell cycle is M phase, or mitosis, in which the duplicated chromosomes separate completely and the cell divides into two daughter cells, each 2n. The two new cells reenter G1 phase and then either continue through another division or cease to divide, entering a quiescent phase (G0) that may last hours, days, or the lifetime of the cell (see Figure 2-10). Differentiated cells such as hepatocytes (liver cells) or adipocytes (fat cells) have acquired their specialized function and thereafter remain in G0 phase.
Many scientists contributed to the description of events during mitosis, and Walther Flemming figures most prominently among them. By the late 1870s, the quality of microscopes included such developments as the oil immersion lens and the substage condenser. These advances made possible Flemming’s detailed observations of dividing cells, published in 1878 and 1882, revealing the stages of mitosis as we know them today. The steps of mitosis are illustrated on the left side of Figure 2-12 and summarized here.
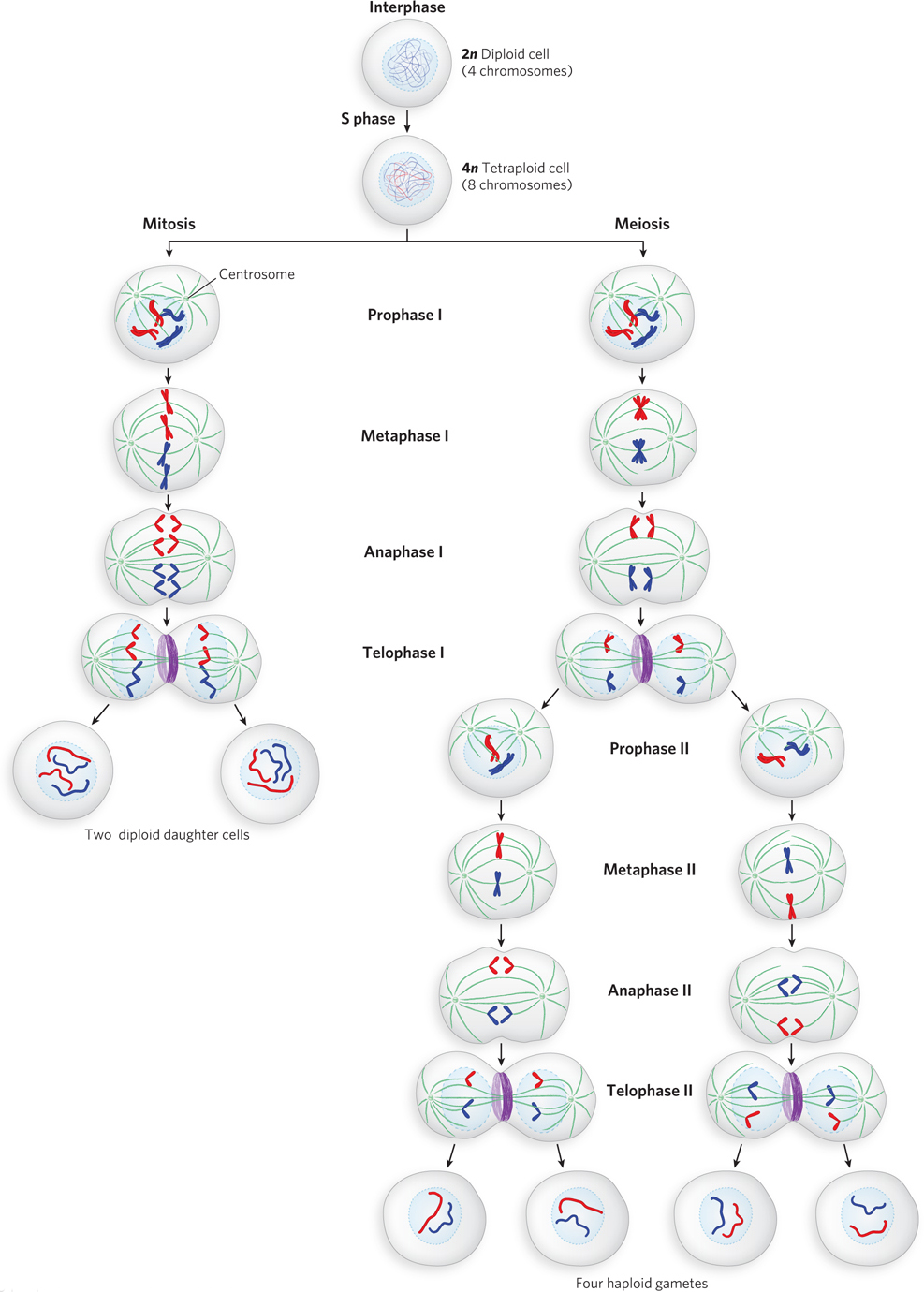
Interphase. Cells not in mitosis are in interphase (which comprises G1, S, and G2). The cell is metabolically active and growing, and the chromosomes are duplicated (in S phase) in preparation for mitosis. The chromosomes are decondensed and not yet visible in the microscope.
Prophase. Prophase is the first stage of mitosis, following G2 phase. As cells enter prophase, the sister chromatid pairs condense and become visible. Two organelles, called centrosomes, move to opposite poles of the cell. There they give rise to the spindle apparatus, an organized structure of protein fibers, which also becomes visible during prophase.
34
35
Metaphase. In metaphase, the membrane surrounding the nucleus dissolves. The spindle apparatus becomes fully developed and attaches to the centromeres of the sister chromatid pairs, directing them to align in the equatorial plane of the cell, a site also known as the metaphase plate.
Anaphase. Each sister chromatid pair separates at the centromere, becoming two separate chromosomes. The spindle apparatus moves the separated chromosomes toward opposite poles of the cell.
Telophase. The two chromosome sets reach opposite cell poles, nuclear membranes (nuclear envelopes) re-
Meiosis: Chromosome Number Is Halved during Gamete Formation
Studies of fertilization in the late 1800s revealed that gamete cells contain only half the number of chromosomes found in somatic cells, and the union of two gametes reestablishes the diploid chromosome number; mitosis keeps this chromosome number constant during somatic cell division. These findings nicely explained how chromosome number is established and maintained in an organism, but they posed a new riddle: how are haploid gamete cells formed? The answer came in the 1880s from studies by Edouard van Beneden, Oskar Hertwig, and Theodor Boveri. Their studies of the ovary cells of a parasitic worm, Ascaris, revealed that the haploid female gamete, the egg (ovum), is formed by two consecutive cell divisions, in a process known as meiosis (see Figure 2-12, right). Later studies revealed that male gametes are also formed by meiotic cell divisions.
The most commonly studied organisms (such as Ascaris, sea urchin, or salamander larvae) had small, similar-
The Process of Meiosis
Meiosis involves a halving of chromosome number to form haploid (n) gametes. One might expect that a diploid cell simply divides once to form two haploid gamete cells, but this is not so. Meiosis involves two successive cell divisions, and four haploid gametes are formed from one diploid cell. The meiotic cell first goes through S phase, just as in mitosis, thereby increasing the number of each chromosome to four copies per cell (4n). In sharp contrast to mitosis, however, the homologous chromosomes—
Whereas mitosis involves only one cell division, the daughter cells from this first meiotic division divide a second time, but without an intervening S phase (no additional chromosome duplication). This second cell division closely resembles mitosis, except that the cells are diploid (2n) going into the second meiotic division (rather than 4n, as in mitosis), so the second division reduces the diploid chromosome number by half, to form haploid gametes (n). In other words, the second meiotic cell division resembles mitosis in that sister chromatids separate, but in meiosis, for each chromosome, there is only one sister chromatid pair to split apart, whereas in mitosis the sister chromatid pairs of both homologous chromosomes are present at the metaphase plate, and each pair splits apart.
The phases of meiosis are summarized (and contrasted with mitosis) here and illustrated in Figure 2-12.
Interphase. Chromosomes are duplicated to form sister chromatid pairs; no obvious difference from mitosis.
Prophase I. Sister chromatid pairs become visible and the spindle apparatus forms. The difference from mitosis is that two homologous sister chromatid pairs find and associate with each other, forming a tetrad. In mitosis, two homologous sister chromatid pairs remain independent and do not associate with each other.
Metaphase I. The nuclear membrane breaks down, and the spindle apparatus moves the four homologous chromosomes to the metaphase plate as a tetrad, rather than moving two homologous but independent sister chromatid pairs as in mitosis.
36
Anaphase I. Centromeres stay intact, and sister chromatids do not separate. Instead, the tetrad splits and the two sets of sister chromatid pairs move to opposite poles. By contrast, in mitosis, sister chromatids split at the centromere and individual chromosomes move apart.
Telophase I. Telophase occurs as in mitosis. The nuclear membrane re-
The second meiotic cell division is a lot like mitosis, but there is no S phase between divisions and the cell is diploid going into the second cell division.
Prophase II. As in mitosis, sister chromatid pairs are visible, but there are half as many as in mitosis because the homologous sister chromatid pair is no longer present (it is in the other daughter cell formed from the first division).
Metaphase II. As in mitosis, the nuclear membrane breaks down and sister chromatid pairs align in the equatorial plane.
Anaphase II. As in mitosis, the centromere splits and the two separated chromosomes move to opposite poles of the cell.
Telophase II. As in mitosis, cytokinesis results in two cells and the two nuclear membranes form. Unlike mitosis, where the resulting daughter cells are diploid (2n), the second meiotic division produces daughter cells that are haploid (n).
In overview, two cells enter meiosis II, and the end result of meiosis is four n cells from a single 2n cell. This is different from mitosis in two ways: the number of daughter cells formed and the n value of each daughter cell.
Sex Determination


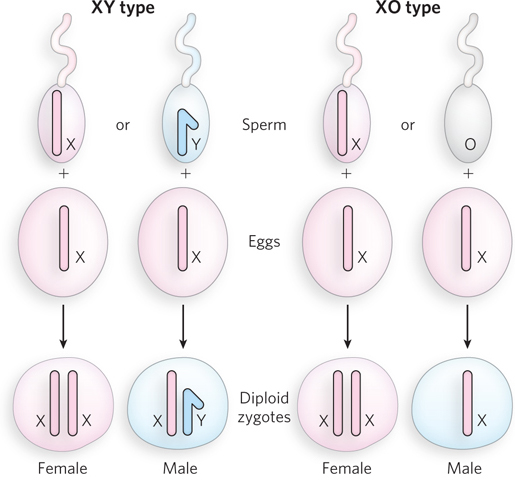
Cytological studies in many types of cells documented the existence of one or two chromosomes that behaved strangely in meiosis during the formation of male gametes. These were called accessory or X chromosomes; they either pair with a morphologically distinct partner chromosome or do not pair at all. Meiotic divisions therefore produce two types of sperm that differ in the accessory chromosome they contain. In 1905, Edmund B. Wilson and Nettie Stevens identified these accessory chromosomes in insects as the determinants of male and female sex and referred to them as X and Y chromosomes, or sex chromosomes. All other chromosomes are called autosomes.
Sex can be determined in many different ways, depending on the type of organism. For example, in mammals, a common way is the XY system (Figure 2-13). In XY determination, the female is XX and the male is XY; the male gametes are of two varieties, carrying either the X or Y chromosome. In many insects, sex is determined by the XO system, in which females are XX and males have one X and no other sex chromosome. The male gametes contain either an X chromosome or no sex chromosome. In both XY and XO determination, the union of male and female gametes is equally likely to produce male or female offspring. In birds, some insects, and other organisms, the ZW system determines sex. It is like the XY system but in reverse: males have two of the same chromosome (ZZ), whereas females have one copy each of the Z and W chromosomes.
37
SECTION 2.2 SUMMARY
Organisms are composed of cells, which have intricate intracellular structures, including chromosomes located in the nucleus.
Cells that are not actively dividing contain two complete sets of unique chromosomes in the nucleus; they are diploid, or 2n (except gametes, which are haploid, or n).
The cell cycle consists of four stages; G1 phase, S phase (synthesis), G2 phase, and M phase (mitosis). The chromosomes of a diploid cell are duplicated in S phase (during interphase) and then carefully segregated into two daughter cells during mitosis, which proceeds through four stages: prophase, metaphase, anaphase, and telophase. The resulting daughter cells are also diploid.
Meiosis is a specialized type of cell division that halves the diploid chromosome number (2n) to produce haploid gametes (n), each containing one complete set of chromosomes.
Haploid gametes unite during fertilization to reestablish the diploid state of the organism.
Sex is determined by an accessory chromosome that is paired either with a similar chromosome or with a distinct, differently shaped chromosome, or has no partner at all. These special chromosomes are called sex chromosomes; all other chromosomes are autosomes.