8.1 How the Eye Works
Life on earth evolved in a world illuminated by the sun during the day and by starlight and the moon’s reflected sunlight at night. Most forms of earthly life are sensitive in one way or another to that light (Land & Furnald, 1992). Even single-celled organisms contain chemicals that respond to light and alter the organism’s activity in survival-promoting ways. In many species of multicellular animals, specialized light-detecting cells called photoreceptors evolved and became connected to the animal’s nervous system. Earthworms, for example, have photoreceptors distributed throughout their skin. Stimulation of these cells by light causes the worm to wriggle back down into the earth, where it finds safety, moisture, and food.
282
1
Through what steps might sophisticated eyes like ours have evolved from primitive beginnings?
Cross-species comparisons, based on homologies (discussed in Chapter 3), suggest that the modern vertebrate eye came about through the following evolutionary steps, or something like them (Gregory, 1996; Lamb et al., 2007): In some early ancestor to vertebrate animals, photoreceptors became concentrated into groups, forming light-detecting organs, or eye spots, just under the skin. These organs may have initially served to enable circadian rhythms—the cyclic biological changes that accompany the day–night light–dark cycle (discussed in Chapter 6). Over successive generations, however, they may have taken on the additional function of responding to shadows, which could be useful in detecting predators. With further natural selection, the skin covering the eyespots became transparent, allowing in both more light and clearer shadows. The spots then gradually moved inward, into fluid-filled pits underneath the transparent skin, which would reduce glare and enable the animal to detect the direction from which changes in illumination were coming. Subsequent evolution led to the thickening of one of the membranes covering each eyespot to form a crude lens, which at first may have served merely to magnify the light reaching the photoreceptors. With further evolutionary refinement, the lens became capable of projecting an image onto the lining of photoreceptors. Through such gradual steps, coupled with appropriate changes in the nervous system, primitive organisms’ ability to detect shifts in lightness and darkness evolved into the ability to see the shapes of things and eventually into the marvelously complex and precise visual ability that is the subject of this chapter.
Functional Organization of the Eye
The main parts of the human eye are shown in Figure 8.1. The photoreceptors lie in the retina, a membrane lining the rear interior of the eyeball. The eyeball is filled with a clear gelatinous substance (the vitreous humor) through which light easily passes. The structures at the front of the eye are devices for focusing light reflected from objects in such a way as to form images on the retina.
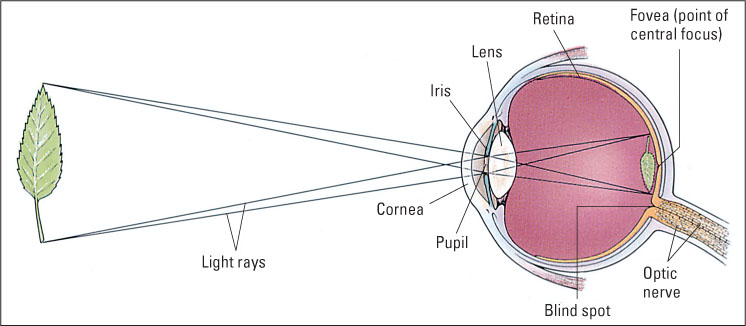
Structures at the Front of the Eye Focus Images on the Retina
2
How do the cornea, iris, and lens help to form images on the retina?
The front of the eyeball is covered by the cornea, a transparent tissue that, because of its convex (outward) curvature, helps focus the light that passes through it. Immediately behind the cornea is the pigmented, doughnut-shaped iris, which provides the color (usually brown or blue) of the eye. The iris is opaque, so the only light that can enter the interior of the eye is that which passes through the pupil, the black-appearing center in the iris, which is simply a hole through which light can pass into the eyeball. Muscle fibers in the iris enable it to increase or decrease the diameter of the pupil to allow more or less light to enter.
Behind the iris is the lens, which adds to the focusing process begun by the cornea. Unlike the cornea, the lens is adjustable; it becomes more spherical when focusing on objects close to the eye and flatter when focusing on those farther away. Light rays diverge as they move toward the eye from any given point on a visual object. The focusing properties of the cornea and lens bring the light rays back together at a particular point on the retina, thereby forming an image of the object on the retina. The image is upside down (as shown at right in Figure 8.1), but that does not matter, since its only purpose is to trigger patterns of activity in neurons running to the brain. The brain is wired to the retina in such a way that input from lower on the retina is interpreted as up, and input from higher on the retina is interpreted as down.
283
Transduction Occurs in the Retina
3
How are cones and rods distributed on the retina, and how do they respond to light?
As noted in Chapter 7, the process by which a stimulus from the environment generates electrical changes in neurons is called transduction. In vision, transduction is the function of the photoreceptor cells. In each eye millions of photoreceptor cells are arranged, mosaic-like, in one thin layer of the multilayered retina. These cells are of two types: cones, which permit sharply focused color vision in bright light, and rods, which permit vision in dim light. They are so named because of their shapes (see Figure 8.2). Cones are most concentrated in the fovea, the pinhead-size area of the retina that is in the most direct line of sight (look again at Figure 8.1), which is specialized for high visual acuity (the ability to distinguish tiny details). The concentration of cones decreases sharply with increasing distance from the fovea. Rods, in contrast, exist everywhere in the retina except in the fovea and are most concentrated in a ring about 20 degrees away from the fovea (see Figure 8.3). Each human retina contains about 6 million cones and 120 million rods (Wade & Swanston, 1991).
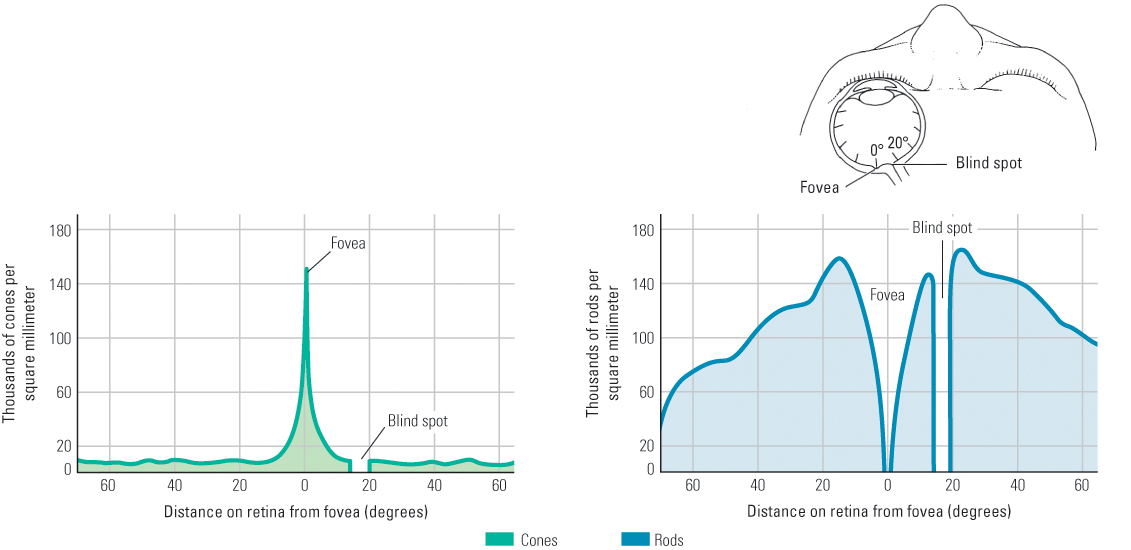
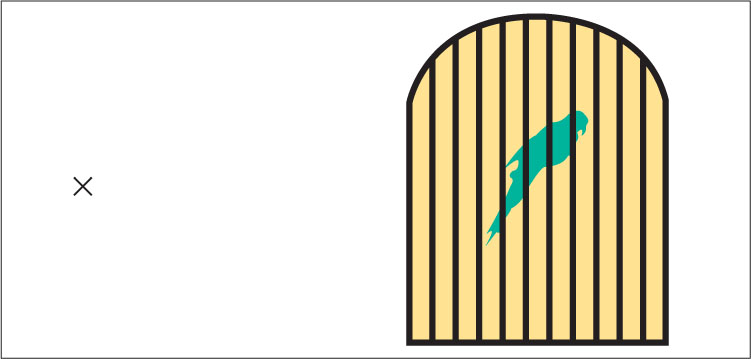
The outer segment of each photoreceptor contains a photochemical—a chemical that reacts to light. The rods’ photochemical is called rhodopsin [rō-dȯp’-sǝn]. When hit by light, rhodopsin molecules undergo a structural change that triggers a series of chemical reactions in the rod’s membrane, which in turn causes a change in the electrical charge across the membrane (Schnapf & Baylor, 1987). The transduction process for cones is similar to that for rods, but (for reasons discussed in the section on color vision) three varieties of cones exist, and each contains a different photochemical. The electrical changes in rods and cones cause electrical responses in other cells in the retina, which lead to the production of action potentials (neural impulses) in neurons that form the optic nerve, which runs from the back of the eye to the brain. At the place on the retina where the axons of these neurons converge to form the optic nerve there is a blind spot, due to the absence of receptor cells (shown in Figure 8.3). We normally do not notice the blind spot, but you can demonstrate its existence by following the instructions in Figure 8.4.
284
Differences Between Cone Vision and Rod Vision
4
How do cone vision and rod vision differ?
Cones and rods provide the starting points for what can be thought of as two separate but interacting visual systems within the human eye. Cone vision, also called photopic vision or bright-light vision, is specialized for high acuity (the ability to see fine detail) and for color perception. Rod vision, also called scotopic vision or dim-light vision, is specialized for sensitivity (the ability to see in very dim light). Rod vision lacks acuity and the capacity to distinguish colors, but, according to calculations from laboratory studies, it is sensitive enough to allow a person on a clear night to detect a single candle flame from 30 miles away if no other lights are present (Galanter, 1962). Cone vision came about, through natural selection, to allow us to see fine details during daylight, and rod vision came about to allow us to see at least the vague outlines of objects at night.
In very dim light, you can see objects best when you don’t look directly at them—because, as noted before, the fovea (the part of the retina in the direct line of sight) contains no rods. You can demonstrate this in a dark room or on a dark night by identifying an object to look at. The object will disappear if you look straight at it, but reappear when you adjust your gaze just a little off to one side.
Roles of Rods and Cones in Dark Adaptation and Light Adaptation
One of the many problems our visual system has to solve is that of adjusting to the enormous range of light intensities that occur over the course of a day. A white object in sunlight at noon reflects roughly 100 million times as much light as the same object reflects on a starlit but moonless night (Riggs, 1965), yet we can see the object in either condition.
5
What is the chemical basis for dark adaptation and light adaptation? Why do we see mostly with cones in bright light and with rods in dim light?
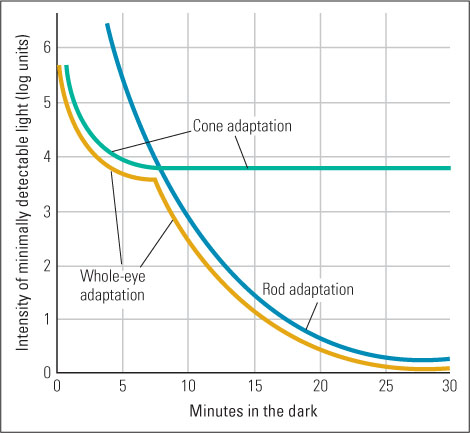
As you know from experience, it takes time for your vision to adapt to sudden large changes in illumination. The gradual increase in sensitivity that occurs after you enter a darkened room is called dark adaptation, and the more rapid decrease in sensitivity that occurs after you turn on a bright lamp or step out into sunlight is called light adaptation. The iris contributes to these adaptive processes by dilating (widening) the pupil in dim light and constricting it in bright light. A fully dilated pupil allows in about 16 times as much light as a fully constricted pupil (Matlin & Foley, 1997). Temporary changes in the sensitivity of visual neurons that receive input from the receptor cells also contribute (Dunn et al., 2007). However, the major contribution to dark and light adaptation comes from the different sensitivities of rods and cones.
285
Rhodopsin, the rod photochemical, is much more sensitive to light than are the cone photochemicals (Kefalov et al., 2003). Bright light causes rhodopsin to break down into two inactive substances, making the rods nonfunctional. In sunlight, therefore, you see entirely with your cones, and even in a typical artificially lit room you see mostly with your cones. When you first step from bright light into a dark room, you may see nothing because your rods are still inactive and there isn’t enough light to activate your cones. After a few moments in the dark, however, rhodopsin begins to regenerate, your rods regain their sensitivity, and you gradually see more and more. It takes about 25 minutes for rhodopsin to regenerate fully. If you then step out of the dark room into sunlight, you are at first blinded by the dazzling light because your highly sensitive rods are responding maximally and indiscriminately. Within about 5 minutes, however, the rhodopsin breaks down and you begin to see normally with your less sensitive cones. The cone photochemicals also break down somewhat in bright light and regenerate in dim light and thereby contribute to light and dark adaptation, but this change is smaller than that which occurs in rods (see Figure 8.5).
SECTION REVIEW
The sophisticated human eye focuses light to form images on photoreceptors in the retina.
Basic Structures and Functions of the Eye
- The cornea and the adjustable lens focus incoming light onto the retina at the back of the eye’s interior.
- Light energy is then transduced by two types of photoreceptor cells in the retina—rods and cones—that contain photochemicals that react to light with structural changes.
- These changes can lead to further chemical and electrical changes, ultimately triggering action potentials in neurons that form the optic nerve.
Cone and Rod Vision
- While cones provide color vision, high visual acuity, and ability to see in bright illumination, rods provide the sensitivity that allows vision in dim illumination.
- The rate and degree of dark adaptation and light adaptation are different for rods and cones. A result is that we see only with rods in very dim light and only with cones in bright light.