8.2 Seeing Colors
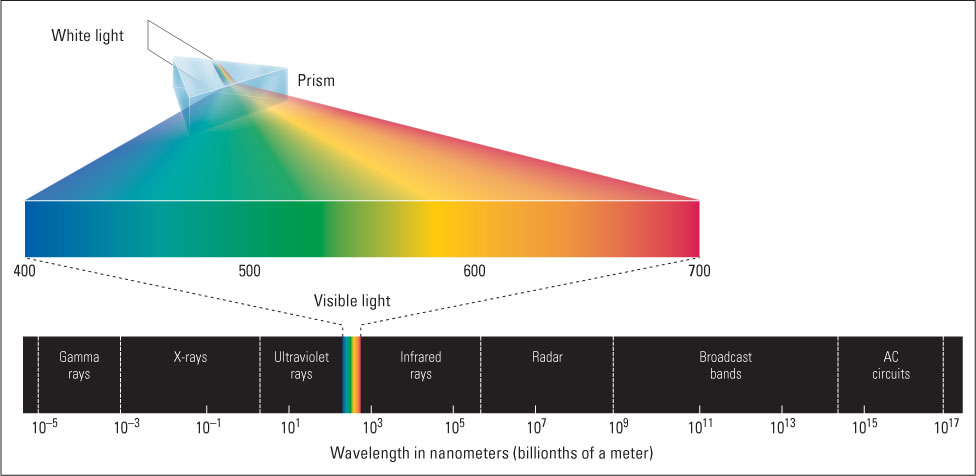
Many animals—including humans, other primates, birds, bees, and most fishes and reptiles—that depend greatly on vision to identify objects have evolved color vision, which helps to make objects stand out vividly from their backgrounds. The colors that we and other animals see in objects depend on the wavelengths of the light that is reflected from those objects. The wavelengths of light visible to humans range from about 400 to 700 nm (1 nm, or nanometer, is a billionth of a meter). Shorter waves of electromagnetic energy, below our visible range, include ultraviolet rays, x-rays, and gamma rays; longer waves, above our visible range, include infrared rays, radar rays, and radio waves (see Figure 8.6).
286
According to the three-primaries law, three different wavelengths of light (called primaries) can be used to match any color that the eye can see if they are mixed in the appropriate proportions. The primaries can be any three wavelengths as long as one is taken from the long-wave end of the spectrum (red), one from the shortwave end (blue or violet), and one from the middle (green or green-yellow). According to the law of complementarity, pairs of wavelengths can be found that, when added together, produce the visual sensation of white. The wavelengths of light in such a pair are referred to as complements of each other.
If you have studied art, you may find this a bit confusing because, although additive color mixing works for light (add all colors of light together and you get white), subtractive color mixing works when dealing with pigments (add all colors of paint together and you get black). When you were a child playing with paint, you probably proved the basic facts of subtractive color mixing many times. You may remember being disappointed when you mixed all the paints together and produced something pretty close to black rather than the brilliant reddish-yellowish-greenish-blue that you had hoped for. In that experiment, you subtracted essentially all the wavelengths by combining all the pigments, leaving essentially none to be reflected. Our discussion of color vision focuses on additive color mixing.
There are two theories of color vision, both developed in the nineteenth century: the trichromatic theory and the opponent-process theory. For many years these two theories were thought to be contradictory to one another, but we now know that both are true.
The Trichromatic Theory
6
How does the trichromatic theory explain the three-primaries law? How was the theory validated by the discovery of three cone types?
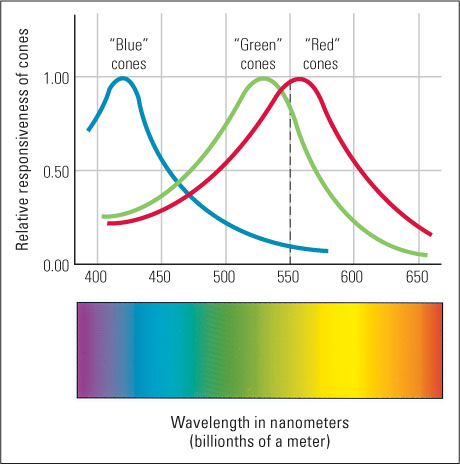
According to the trichromatic theory, color vision emerges from the combined activity of three different types of receptors, each most sensitive to a different range of wavelengths. This idea was first proposed (in 1802) by Thomas Young and later by Hermann von Helmholtz (1852). Young and Helmholtz reasoned that if every color we see is the result of a unique proportion, or ratio, of activity among three types of receptors, then the three-primaries law would be an inevitable result: It would be possible to match any visible color by varying the relative intensities of three primary lights, each of which acts maximally on a different type of receptor. Young and Helmholtz developed their theory purely from behavioral data on the perceptual effects of color mixing, at a time when nothing was known about photoreceptors in the retina. We now know from physiological studies that their theory was correct. Three types of cones indeed exist in the human retina, each with a different photochemical that makes it most sensitive to the light within a particular band of wavelengths.
287
In Figure 8.7 you can see an approximation of the actual sensitivity curves for each type of cone. The cones are labeled “blue,” “green,” and “red,” after the color that is experienced when that type of cone is much more active than the other types. Notice that any given wavelength of light produces a unique ratio of activity in the three types of cones. For example, a 550-nm light, which is seen as greenish-yellow, produces a slightly larger response in “red” cones than in “green” cones and a very low response in “blue” cones. That same ratio of response in the three cone types could be produced by shining into the eye a mixture of red, green, and blue primaries, with the first two much more intense than the last. The result would be a perceptual experience of greenish-yellow indistinguishable from that produced by the 550-nm light.
Exceptions to Trichromatic Vision
7
Why does vision in some people obey a two-primaries law rather than the three-primaries law, and why are these people not good at picking cherries? How does the color vision of most nonprimate mammals, and that of most birds, differ from that of most humans?
Some people (one of your authors [David Bjorklund] being one of them), referred to as dichromats, have only two, not three, types of cone photochemicals. These people, as you might expect, obey a two-primaries law of color mixing rather than the three-primaries law. For them, any color that they can see can be matched by varying the proportion of just two different wavelengths of light (see Figure 8.7).
The most common forms of dichromia involve the absence of the normal photochemical for either the “red” or the “green” cones (usually the latter) due to a defect in the gene that produces that photochemical (Neitz et al., 1996). Because the defective gene is recessive and the genes for both the “red” and the “green” photochemicals are located on the X chromosome, this trait appears much more often in men than in women. Men (as discussed in Chapter 3) have only one X chromosome, inherited from the mother, so a single defective gene on that chromosome can produce color blindness (see Figure 8.8).

Garo/Phanie/Science Source
288
Approximately 5 percent of men and 0.25 percent of women lack either the “red” or “green” cone photochemical and are red-green color-blind, meaning that they have difficulty distinguishing colors ranging from green to the red end of the spectrum (Masland, 2001). If you look again at Figure 8.7, you will see why this is so. The normal ability to distinguish colors in this range (from about 500 to 700 nm) is mediated almost entirely by differential activity in the “red” and “green” cones, because “blue” cones are quite inactive in this range. If either the “red” or the “green” cone photochemical is missing, then the person has no or little physiological basis for distinguishing one wavelength from another in this range.
Many people who have red-green color blindness don’t know it and may wonder why certain perceptual tasks that are hard for them are easy for others. One man’s red-green color blindness was not discovered until he told his family how much he admired the perceptual skill of cherry pickers: “After all,” he said, “the only thing that tells ’em it’s a cherry is … that it’s round and the leaves aren’t. I just don’t see how they find ’em in those trees!” (Coren & Ward, 1989). There is an upside to some forms of color blindness, however. Color-blind people are not easily fooled by camouflage; in World War II the U.S. Army even assigned color-blind personnel to detect camouflage colors that people with normal color vision could not see (Reit, 1978).

© David R. Frazier/The Image Works
Most nonprimate mammals, including dogs and cats, have just two types of cones, corresponding to our “blue” and “green” cones, and have the same difficulty discriminating among colors at the long-wavelength end of the spectrum as do people who are red-green color-blind (Solomon & Lennie, 2007). Most birds, in contrast, have four types of cones (Dalton, 2004). Birds rely heavily on vision to guide their flight and to locate food, and they have evolved color vision that is better than ours. In addition to “red,” “green,” and “blue” cones, they have a fourth set of cones that are maximally sensitive to wavelengths in the ultraviolet range (shorter than 400 nm), which we cannot see but are visible to other animals, such as bird species whose hatchlings have ultraviolet mouths.
289
The Opponent-Process Theory

The trichromatic theory explains the three-primaries law and certain types of color blindness well, but it does not explain the law of complementarity—how certain pairs of wavelengths produce the experience of white. To explain that, Ewald Hering, another nineteenth-century scientist, developed the opponent-process theory. Hering was most impressed by the observation that complementary colors of light (blue and yellow, or green and red) seem to swallow each other up, erasing each other’s color, when added together. For example, if you begin with blue light and gradually add more of its complement (yellow), the result is not “bluish-yellow” but an ever-paler (more unsaturated) blue, which finally becomes white. To explain such observations, Hering (1878/1964) proposed that color perception is mediated by physiological units (which we now call neurons) that can be either excited or inhibited, depending on the wavelength of light, and that complementary wavelengths have opposite effects (that is, they activate “opposing processes”) on these opponent-process units.
8
How does the opponent-process theory explain (a) the law of complementarity in color mixing and (b) the complementarity of afterimages?
More specifically, Hering’s proposal (in modern language) was that the ability to see blues and yellows is mediated by blue-yellow opponent neurons, which are excited by wavelengths in the blue part of the spectrum and inhibited by those in the yellow part, or vice versa. Similarly, he proposed that the ability to see greens and reds is mediated by green-red opponent neurons, which are excited by wavelengths in the green part of the spectrum and inhibited by those in the red part, or vice versa. In addition, he proposed that the ability to distinguish bright from dim light, independent of wavelength, is mediated by a third set of neurons (brightness detectors), which are excited by lights of any wavelength. This theory nicely accounts for the facts of complementary colors. A mixture of wavelengths from the blue and yellow parts of the spectrum, or from the green and red parts, appears white (colorless but bright) because the two sets of wavelengths cancel each other out in their effects on color detectors but act in concert to excite brightness detectors.
Color Afterimages Explained by the Opponent-Process Theory
The opponent-process theory also accounts wonderfully for another psychological phenomenon, the complementarity of afterimages. For a demonstration of this phenomenon, follow the instructions in the first paragraph of the caption of Figure 8.9. You will see that the colors in the afterimage are the complements of those in the original: What was green becomes red; what was yellow becomes blue; and what was black becomes white.

To demonstrate that this effect involves a change within the retina, or at least somewhere early in the visual system before the input from the two eyes converges, repeat the demonstration but this time keep one eye closed as you stare at the middle of the flag. Then look at the dot in the white space first with one eye and then with the other. What happens?
290
How does the opponent-process theory explain this phenomenon? Consider the example of green becoming red in the afterimage. The neurons in the retina that respond most strongly to the green-appearing (middle-wavelength) light as you stare at the picture become fatigued. Therefore, when you shift your eyes to the white paper (which reflects all wavelengths), those neurons don’t respond as strongly as they normally would, but other neurons, including those that respond to red-appearing (long-wavelength) light, do respond strongly. Thus, opponent-process neurons that are normally excited by red-appearing light and inhibited by green-appearing light in that part of the retina become excited, resulting in the perception of red. To convince yourself that this adaptive process occurs early in the visual pathway, before the inputs from the two eyes converge, follow the instructions in the second paragraph of the caption of Figure 8.9.
A Physiological Reconciliation of the Two Theories
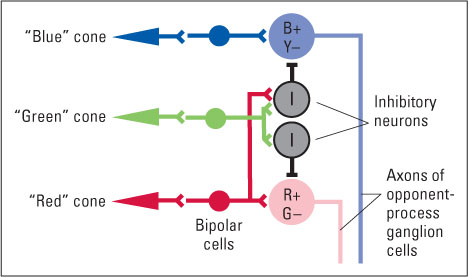
For many years the trichromatic and opponent-process theories were thought to be contradictory, but in the 1950s and 1960s research showed that both theories are fundamentally correct (De Valois et al., 1966; Hurvich & Jameson, 1957). The retina indeed contains three types of cones, consistent with Young and Helmholtz’s trichromatic theory. But the cones feed into ganglion cells (the neurons of the optic nerve) in a pattern that translates the trichromatic code into an opponent-process code, conforming to Hering’s theory. Some ganglion cells behave in a red-green opponent manner: They are excited by input from “red” cones and inhibited by input from “green” cones, or vice versa. Others behave in a blue-yellow opponent manner: They are excited by input from “blue” cones and inhibited by input from both “green” and “red” cones (which, in combination, respond best to yellow light), or vice versa. To see how connections from the three cone types could produce such opponent-process cells, look at Figure 8.10. Neurons in the cerebral cortex that are most directly involved in color perception maintain these opponent-process characteristics (Dacey, 2000; Solomon & Lennie, 2007).
9
How has the opponent-process theory been validated in studies of the activity of neurons that receive input from cones?
The research and theories on color vision just presented are a wonderful early illustration of the value of combining behavioral and physiological research. The trichromatic and opponent-process theories were developed, in the nineteenth century, from behavioral evidence having to do with the perceptual effects of color mixing, before anything was known about the physiology of receptors and neurons. Later, both theories were confirmed physiologically, and today neuroscientists and psychologists are working out the finer details of the theories.
SECTION REVIEW
The experience of color results from the visual system’s response to wavelengths of light.
Wavelength and Color
- We experience different wavelengths of light as different colors.
- Objects appear colored because their pigments absorb some wavelengths from white light and reflect others.
- Color mixing involves mixing lights of different wavelengths. It is characterized by the three-primaries law and the law of complementarity.
Trichromatic Theory of Color Vision
- This theory, derived from the three-primaries law, holds that we have three types of receptors, each most responsive to a particular range of wavelengths.
- The theory was confirmed by the discovery that the retina has three types of cones, each with a different curve of sensitivity to visible wavelengths.
Opponent-Process Theory of Color Vision
- This theory, derived from the law of complementarity, holds that physiological units involved in color vision are affected in opposite ways (excited or inhibited) by complementary wavelengths.
- The theory explains the complementarity of afterimages.
- The theory was confirmed by the discovery of visual neurons that behave just as the theory predicted.
291