CONCEPT18.3 Major Events in the Evolution of Life Can Be Read in the Fossil Record
How do we know about the physical changes in Earth’s environment and their effects on the evolution of life? To reconstruct life’s history, scientists rely heavily on the fossil record. As we have seen, geologists divided Earth’s history into eons, eras, and periods based on their distinct fossil assemblages (see Table 18.1). Biologists refer to the assemblage of all organisms of all kinds living at a particular time or place as a biota. All of the plants living at a particular time or place are its flora, and all of the animals are its fauna.
About 300,000 species of fossil organisms have been described, and the number steadily grows. The number of named species, however, is only a tiny fraction of the species that have ever lived. We do not know how many species lived in the past, but we have ways of making reasonable estimates. Of the present-day biota, nearly 1.8 million species have been named. The actual number of living species is probably well over 10 million, and possibly much higher, because many species have not yet been discovered and described by biologists. So the number of described fossil species is only about 3 percent of the estimated minimum number of living species. Life has existed on Earth for about 3.8 billion years. Many species last only a few million years before undergoing speciation or going extinct. From this we know that Earth’s biota must have turned over many times during geological history. So the total number of species that have lived over evolutionary time must vastly exceed the number living today. Why have only about 300,000 of these tens of millions of species been described from fossils to date?
Several processes contribute to the paucity of fossils
Only a tiny fraction of organisms ever become fossils, and only a tiny fraction of fossils are ever discovered by paleontologists. Most organisms live and die in oxygen-rich environments in which they quickly decompose. Organisms are not likely to become fossils unless they are transported by wind or water to sites that lack oxygen, where decomposition proceeds slowly or not at all. Furthermore, geological processes transform many rocks, destroying the fossils they contain, and many fossil-bearing rocks are deeply buried and inaccessible. Paleontologists have studied only a tiny fraction of the sites that contain fossils, although they find and describe many new ones every year.
The fossil record is most complete for marine animals that had hard skeletons (which resist decomposition). Among the nine major animal groups with hard-shelled members, approximately 200,000 species have been described from fossils—roughly twice the number of living marine species in these same groups. Paleontologists lean heavily on these groups in their interpretations of the evolution of life. Insects and spiders are also relatively well represented in the fossil record because they are numerically abundant and have hard exoskeletons (FIGURE 18.9). The fossil record, though incomplete, is good enough to document clearly the factual history of the evolution of life.

By combining information about physical changes during Earth’s history with evidence from the fossil record, scientists have composed portraits of what Earth and its inhabitants may have looked like at different times. We know in general where the continents were and how life changed over time, but many of the details are poorly known, especially for events in the more remote past.
Precambrian life was small and aquatic
Life first appeared on Earth about 3.8 bya (FIGURE 18.10). The fossil record of organisms that lived prior to the Phanerozoic is fragmentary, but it is good enough to establish that the total number of species and individuals increased dramatically in the late Precambrian.
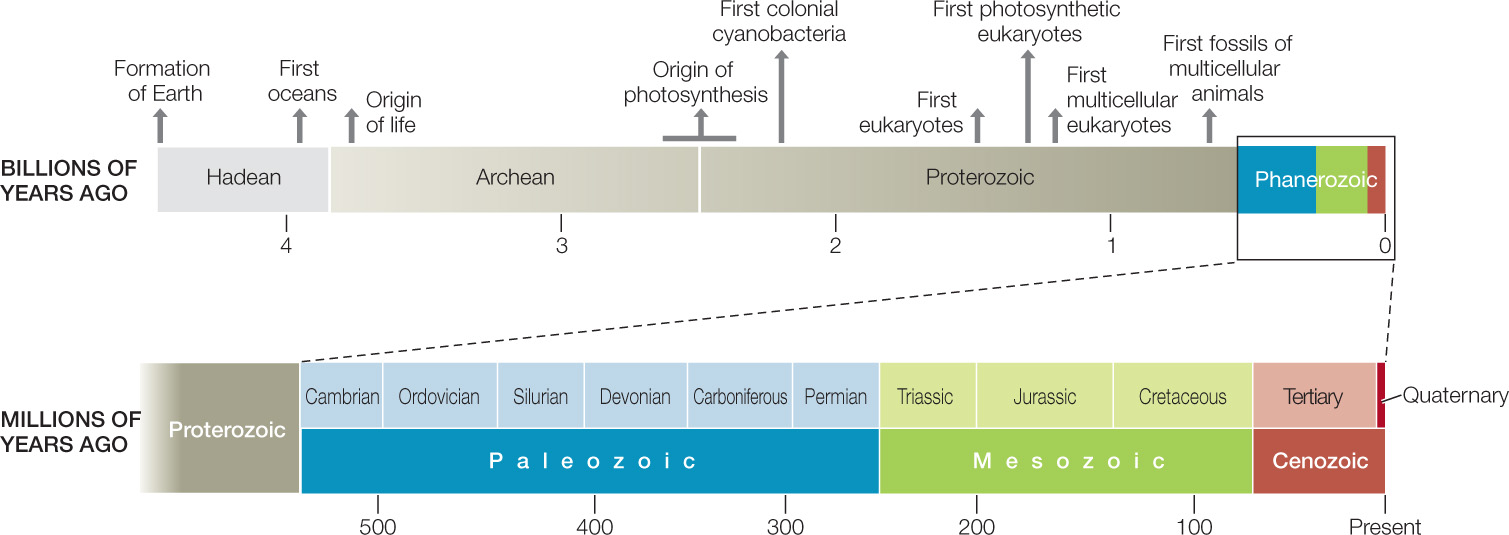
For most of its history, life was confined to the oceans, and all organisms were small. For more than 3 billion years, all organisms lived in shallow seas. These seas slowly began to teem with microscopic prokaryotes. After the first eukaryotes appeared about 1.5 billion years ago, during the Proterozoic, unicellular eukaryotes and small multicellular animals fed on the microorganisms. Small floating organisms, known collectively as plankton, were strained from the water and eaten by slightly larger filter-feeding animals. Other animals ingested sediments on the seafloor and digested the remains of organisms within them. But it still took nearly a billion years before eukaryotes began to diversify rapidly into the many different morphological forms that we know today.
What limited the diversity of multicellular eukaryotes (in terms of their size and shape) for much of their early existence? It is likely that a combination of factors was responsible. We have already noted that O2 levels increased throughout the Proterozoic, and it is likely that high atmospheric and dissolved O2 concentrations were needed to support large multicellular organisms. In addition, geologic evidence points to a series of intensely cold periods during the late Proterozoic, which would have resulted in seas that were largely covered by ice and continents that were covered by glaciers. The “snowball Earth” hypothesis suggests that cold conditions confined life to warm places such as hot springs, deep thermal vents, and perhaps a few equatorial oceans that avoided ice cover. The last of these Proterozoic glaciations ended about 580 million years ago, just before several major radiations of multicellular eukaryotes appear in the fossil record (FIGURE 18.11). Many of the multicellular organisms known from the late Proterozoic and early Phanerozoic were very different from any animals living today and may be members of groups that left no living descendants.
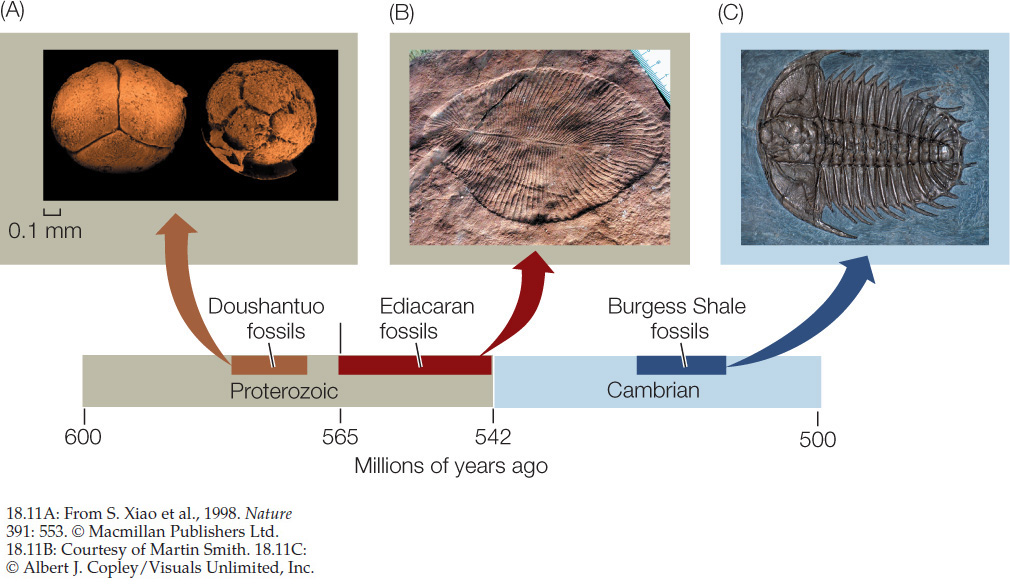
Life expanded rapidly during the Cambrian period
The Cambrian period (542–488 mya) marks the beginning of the Paleozoic, the first era of the Phanerozoic. The O2 concentration in the Cambrian atmosphere was approaching its current level, and the glaciations of the late Proterozoic had ended nearly 40 million years earlier. A geologically rapid diversification of life took place that is often called the Cambrian explosion. This name is somewhat misleading, as the series of radiations it refers to actually began before the start of the Cambrian and continued for about 60 million years into the early Cambrian (see Figure 18.11). Nonetheless, 60 million years represents a relatively short amount of time, especially considering that the first eukaryotes had appeared about a billion (= 1,000 million) years earlier. Many of the major animal groups represented by species alive today first appeared during these evolutionary radiations. FIGURE 18.12 provides an overview of the numerous continental and biotic innovations that have characterized the Phanerozoic.

For the most part, fossils tell us only about the hard parts of organisms, but in some well-studied Cambrian fossil beds, the soft parts of many animals were preserved. Multicellular life was largely or completely aquatic during the Cambrian. If there was life on land at this time, it was probably restricted to microorganisms.
Many groups of organisms that arose during the Cambrian later diversified
Geologists divide the remainder of the Paleozoic era into the Ordovician, Silurian, Devonian, Carboniferous, and Permian periods. Each period is characterized by the diversification of specific groups of organisms. Mass extinctions marked the ends of the Ordovician, Devonian, and Permian.
The Ordovician (488-444 MYA)
During the Ordovician period, the continents, which were located primarily in the Southern Hemisphere, still lacked multicellular life. Evolutionary radiation of marine organisms was spectacular during the early Ordovician, especially among animals, such as brachiopods and mollusks, that lived on the seafloor and filtered small prey from the water. At the end of the Ordovician, as massive glaciers formed over the southern continents, sea levels dropped about 50 meters, and ocean temperatures dropped. About 75 percent of all animal species became extinct, probably because of these major environmental changes.
The Silurian (444-416 MYA)
During the Silurian period, the continents began to merge together. Marine life rebounded from the mass extinction at the end of the Ordovician. Animals able to swim in open water and feed above the ocean floor appeared for the first time. Fishes diversified as bony armor gave way to the less rigid scales of modern fishes, and the first jawed fishes and the first fishes with supporting rays in their fins appeared. The tropical sea was uninterrupted by land barriers, and most marine organisms were widely distributed. On land, the first vascular plants evolved late in the Silurian (about 420 mya). The first terrestrial arthropods—scorpions and millipedes—evolved at about the same time.
The Devonian (416-359 MYA)
Rates of evolutionary change accelerated in many groups of organisms during the Devonian period. The major land masses continued to move slowly toward each other. In the oceans there were great evolutionary radiations of corals and of shelled, squidlike cephalopod mollusks.
Terrestrial communities changed dramatically during the Devonian. Club mosses, horsetails, and tree ferns became common, and some attained the size of large trees. Their roots accelerated the weathering of rocks, resulting in the development of the first forest soils. The first plants to produce seeds appeared in the Devonian. The earliest fossil centipedes, spiders, mites, and insects date to this period, as do the earliest terrestrial vertebrates.
A massive extinction of about 75 percent of all marine species marked the end of the Devonian. Paleontologists are uncertain about its cause, but two large meteorites collided with Earth at about that time and may have been responsible, or at least a contributing factor. The continued merging of the continents, with the corresponding reduction in the area of continental shelves, also may have contributed to this mass extinction.
The Carboniferous (359-299 MYA)
Large glaciers formed over high-latitude portions of the southern land masses during the Carboniferous period, but extensive swamp forests grew on the tropical continents. These forests were dominated by giant tree ferns and horsetails with small leaves. Their fossilized remains formed the coal we now mine for energy. In the seas, crinoids (a group of echinoderms, related to sea stars and sea urchins) reached their greatest diversity, forming “meadows” on the seafloor.
The diversity of terrestrial animals increased greatly during the Carboniferous. Snails, scorpions, centipedes, and insects were abundant and diverse. Insects evolved wings, becoming the first animals to fly. Flight gave herbivorous insects easy access to tall plants, and plant fossils from this period show evidence of chewing by insects (FIGURE 18.13). The terrestrial vertebrates split into two lineages. The amphibians became larger and better adapted to terrestrial existence, while the sister lineage led to the amniotes—vertebrates with well-protected eggs that can be laid in dry places.

The Permian (299-251 MYA)
During the Permian period, the continents merged into a single supercontinent called Pangaea. Permian rocks contain representatives of many of the major groups of insects we know today. By the end of the period the amniotes had split into two lineages: the reptiles, and a second lineage that would lead to the mammals. Ray-finned fishes became common in the freshwaters of Pangaea.
Toward the end of the Permian, conditions for life deteriorated. Massive volcanic eruptions resulted in outpourings of lava that covered large areas of Earth. The ash and gases produced by the volcanoes blocked sunlight and cooled the climate. The death and decay of the massive Permian forests rapidly used up atmospheric oxygen, and the loss of photosynthetic organisms meant that relatively little new atmospheric oxygen was produced. In addition, much of Pangaea was located close to the South Pole by the end of the Permian. All of these factors combined to produce the most extensive continental glaciers since the “snowball Earth” times of the late Proterozoic. Atmospheric oxygen concentrations gradually dropped from about 30 percent to 15 percent. At such low concentrations, most animals would have been unable to survive at elevations above 500 meters, so about half of the land area would have been uninhabitable at the end of the Permian. The combination of these changes resulted in the most drastic mass extinction in Earth’s history. Scientists estimate that about 96 percent of all multicellular species became extinct at the end of the Permian.
Geographic differentiation increased during the Mesozoic era
The few organisms that survived the Permian mass extinction found themselves in a relatively empty world at the start of the Mesozoic era (251 mya). As Pangaea slowly began to break apart in the Mesozoic, the biotas of the newly separated continents began to diverge. The oceans rose and once again flooded the continental shelves, forming huge, shallow inland seas. Atmospheric oxygen concentrations gradually rose. Life once again proliferated and diversified, but different groups of organisms came to the fore. The three groups of phytoplankton (floating photosynthetic organisms) that dominate today’s oceans—dinoflagellates, coccolithophores, and diatoms—became ecologically important at this time, and their remains are the primary origin of the world’s oil deposits. Seed-bearing plants replaced the trees that had ruled the Permian forests.
The Mesozoic era is divided into three periods: the Triassic, Jurassic, and Cretaceous. The Triassic and Cretaceous were terminated by mass extinctions, probably caused by meteorite impacts.
The Triassic (251-201.6 MYA)
Pangaea remained largely intact through the Triassic. Many invertebrate groups diversified, and many burrowing animals evolved from groups living on the surfaces of seafloor sediments. On land, conifers and seed ferns were the dominant trees. The first frogs and turtles appeared. A great radiation of reptiles began, which eventually gave rise to crocodilians, dinosaurs, and birds. The first mammals appear. The end of the Triassic was marked by a mass extinction that eliminated about 65 percent of the species on Earth.
The Jurassic (201.6-145.5 MYA)
Late in the Jurassic period, Pangaea became fully divided into two large continents: Laurasia, which drifted northward, and Gondwana in the south. Ray-finned fishes rapidly diversified in the oceans. The first lizards appeared, and flying reptiles (pterosaurs) evolved. Most of the large terrestrial predators and herbivores of the period were dinosaurs. Several groups of mammals made their first appearance, and the earliest known fossils of flowering plants are from late in this period.
The Cretaceous (145.5-65.5 MYA)
By the mid-Cretaceous period, Laurasia and Gondwana had largely broken apart into the continents we know today (although the Indian subcontinent was still separated from Asia). A continuous sea encircled the tropics. Sea levels were high, and Earth was warm and humid. Life proliferated both on land and in the oceans. Marine invertebrates increased in diversity. On land, the reptile radiation continued as dinosaurs diversified further and the first snakes appeared. Early in the Cretaceous, flowering plants began the radiation that led to their current dominance of the land. By the end of the period, many groups of mammals had appeared.
As described in Concept 18.2, another meteorite-caused mass extinction took place at the end of the Cretaceous. In the seas, many planktonic organisms and bottom-dwelling invertebrates became extinct. On land, almost all animals larger than about 25 kg in body weight became extinct. Many species of insects died out, perhaps because the growth of the plants they fed upon was greatly reduced following the impact. Some species in northern North America and Eurasia survived in areas that were not subjected to the devastating fires that engulfed most low-latitude regions.
Modern biotas evolved during the Cenozoic era
By the early Cenozoic era (65.5 mya), the continents were getting closer to their present positions, but the Indian subcontinent was still separated from Asia, and the Atlantic Ocean was much narrower. The Cenozoic was characterized by an extensive radiation of mammals, but other groups were also undergoing important changes.
Flowering plants diversified extensively and came to dominate world forests, except in the coolest regions, where the forests were composed primarily of gymnosperms. Mutations of two genes in one group of plants (the legumes) allowed them to use atmospheric nitrogen directly by forming symbioses with a few species of nitrogen-fixing bacteria. The evolution of this symbiosis was the first “green revolution” and dramatically increased the amount of nitrogen available for terrestrial plant growth. This symbiosis remains fundamental to the ecological base of life as we know it today.
LINK
The symbiosis between plants and nitrogen-fixing bacteria is covered in detail in Concept 25.2
The Cenozoic era is divided into the Tertiary and the Quaternary periods, which are commonly subdivided into epochs (TABLE 18.2).
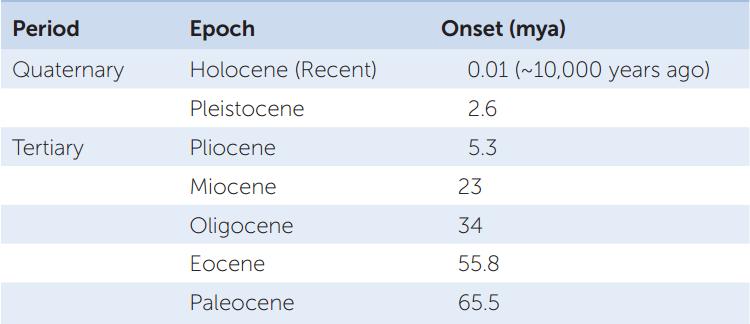
The Tertiary (65.5-2.6 MYA)
During the Tertiary period, the Indian subcontinent continued its northward drift. By about 55 mya it made initial contact with parts of southeastern Asia. By about 35 mya, the Indian Plate ran fully into the Eurasian Plate, and the Himalayas began to be pushed up as a result.
The early Tertiary was a hot and humid time, and the ranges of many plants shifted latitudinally. The tropics were probably too hot to support rainforest vegetation and instead were clothed in low-lying vegetation. In the middle of the Tertiary, however, Earth’s climate became considerably cooler and drier. Many lineages of flowering plants evolved herbaceous (non-woody) forms, and grasslands spread over much of Earth.
By the start of the Cenozoic era, invertebrate faunas had already come to resemble those of today. Frogs, snakes, lizards, birds, and mammals all underwent extensive radiations during the Tertiary. Three waves of mammals dispersed from Asia to North America across one of the several land bridges that have intermittently connected the two continents during the past 55 million years. Rodents, marsupials, primates, and hoofed mammals appeared in North America for the first time.
The Quaternary (2.6 MYA TO PRESENT)
We are living in the Quaternary period. It is subdivided into two epochs, the Pleistocene and the Holocene (the Holocene is also known as the Recent).
The Pleistocene was a time of drastic cooling and climate fluctuations. During 4 major and about 20 minor “ice ages,” massive glaciers spread across the continents, and the ranges of animal and plant populations shifted toward the equator. The last of these glaciers retreated from temperate latitudes less than 15,000 years ago. Organisms are still adjusting to this change. Many high-latitude ecological communities have occupied their current locations for no more than a few thousand years.
It was during the Pleistocene that divergence within one group of mammals, the primates, resulted in the evolution of the hominoid lineage. Subsequent hominoid radiation eventually led to the species Homo sapiens—modern humans. Many large bird and mammal species became extinct in Australia and in the Americas when H. sapiens arrived on those continents about 45,000 and 15,000 years ago, respectively. Many paleontologists believe these extinctions were the result of hunting and other influences of Homo sapiens.
LINK
The evolution of modern humans and their close relatives during the Pleistocene is discussed in Concept 23.7
The tree of life is used to reconstruct evolutionary events
The fossil record reveals broad patterns in life’s evolution. To reconstruct major events in the history of life, biologists also rely on the phylogenetic information in the tree of life. We can use phylogeny, in combination with the fossil record, to reconstruct the timing of such major events as the acquisition of mitochondria in the ancestral eukaryotic cell, the several independent origins of multicellular organisms, and the movement of life onto dry land. We can also follow major changes in the genomes of organisms, and we can even reconstruct many gene sequences of species that are long extinct.
LINK
Concept 16.3 describes how biologists reconstruct the gene sequences of extinct organisms
Changes in Earth’s physical environment have clearly influenced the diversity of organisms we see on the planet today. To study the evolution of that diversity, biologists examine the evolutionary relationships among species. Deciphering phylogenetic relationships is an important step in understanding how life has diversified on Earth. The next part of this book will explore the major groups of life and the different solutions these groups have evolved to major challenges such as reproduction, energy acquisition, dispersal, and escape from predation.
CHECKpointCONCEPT18.3
- Why have so few of the organisms that have existed over Earth’s history become fossilized?
- What do we mean by the “Cambrian explosion”? How long did it last, and in what sense was it an “explosion”?
- What are some of the ways in which continental drift has affected the evolution of life?
Can modern experiments test hypotheses about the evolutionary impact of ancient environmental changes?
ANSWER Several experiments have been conducted to test the link between O2 concentrations and evolution of body size in flying insects. One of these is discussed in Figure 18.8. The results of these experiments are consistent with the evolution of larger body size in flying insects in hyperoxic (high-oxygen) environments.
Experiments have also been conducted under hypoxic (low-oxygen) conditions, as existed at the end of the Permian. These experiments suggest that the evolution of body size is constrained under hypoxic conditions, even under strong artificial selection for larger body size. These latter results are consistent with the extinction of many large flying insects at the end of the Permian as a result of rapidly decreasing O2 concentrations. Giant flying insects simply could not have survived the lower O2 concentrations that existed at that time. The mass extinction at the end of the Permian is the only known mass extinction that involved considerable loss of insect diversity.