CONCEPT33.4 Many Distinctive Types of Muscle Have Evolved
Besides skeletal muscle, several other distinctive types of muscle have evolved. Vertebrate cardiac muscle—heart muscle—is one type. During each of our lives, our cardiac muscle beats regularly from our early embryonic development until we die.
Vertebrate cardiac muscle is both similar to and different from skeletal muscle
Like skeletal muscle, cardiac muscle appears striated when viewed under a light microscope. This is because the actin and myosin filaments in cardiac muscle are arranged in regular patterns in sarcomeres. In other ways, however, cardiac muscle differs morphologically from skeletal muscle. Cardiac muscle cells are much smaller than skeletal muscle cells, and unlike skeletal muscle cells, they have only one nucleus.
A key way that cardiac muscle differs from skeletal muscle is in the relationship of adjacent cells. In cardiac muscle, adjacent cells are electrically coupled by gap junctions that occur in structures called intercalated discs. At a gap junction, the sarcoplasms (cytoplasms) of two adjacent cardiac muscle cells are continuous with each other via tiny pores:
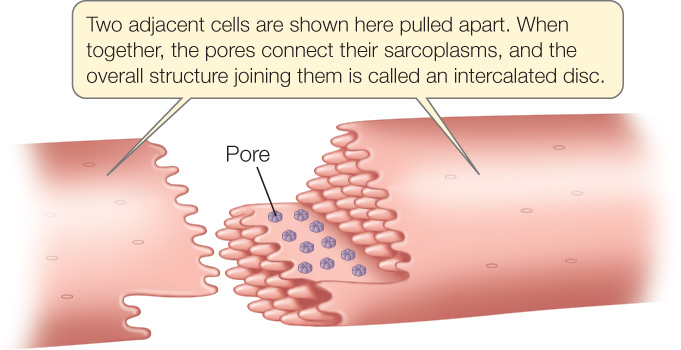
Because of this continuity, if one of the two cells is excited and an impulse spreads over its cell membrane, the impulse spreads directly to the second cell (see Figure 4.18 and Concept 4.5). Accordingly, when an impulse is initiated at any one point in a mass of heart muscle, it spreads rapidly throughout the muscle by moving directly from one cell to another. This process ensures that all the cells within a large mass of muscle are excited at approximately the same time. Thus all the cells contract almost simultaneously, raising the blood pressure in the heart to a high level and propelling the blood out of the heart and through the blood vessels.
In a vertebrate heart, certain of the cells in the heart muscle itself generate the heartbeat rhythm (see Concept 32.3 and Figure 32.8). The heart rhythm is thus very different from the breathing rhythm. The breathing muscles of vertebrates are skeletal muscles, and like all skeletal muscles, they contract only if stimulated by nerves. If a person suffers a neck fracture, he or she will stop breathing if the nerves that stimulate the breathing muscles are damaged. His or her heart will keep beating, however, because of the properties of certain of the cardiac muscle cells.
Vertebrate smooth muscle powers slow contractions of many internal organs
In addition to skeletal and cardiac muscle, vertebrate animals have a third type of muscle known as smooth muscle. Structurally, smooth muscle cells are smaller than skeletal muscle cells. They are usually spindle-shaped, and each has a single nucleus. They are termed “smooth” because their actin and myosin filaments are not as regularly arranged as those in skeletal and cardiac muscle, and for this reason they do not have a striated appearance when viewed under a light microscope:
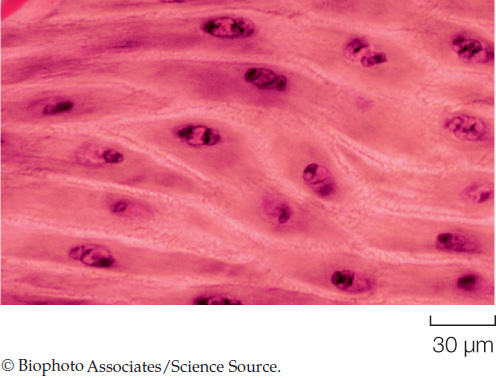
Smooth muscles provide the contractile forces for most of our internal organs, under the control of the nervous system. Smooth muscles in the walls of the digestive tract move food through the digestive tract. Smooth muscles in the walls of small arteries and arterioles adjust the diameters of these blood vessels, helping control blood flow through them. During sexual climax, smooth muscles in the ejaculatory passages of the male propel the ejaculation of semen, and in the female smooth muscles contribute to the rhythmic contractions of orgasm. Smooth muscles in the walls of the bladder produce forces for bladder emptying.
Some smooth muscle tissue, such as that in the walls of the digestive tract, consists of sheets of cells in which adjacent cells are in electrical contact through gap junctions. As a result, excitation of one cell can spread directly to adjacent cells, helping coordinate the contractions of the cells. This occurs during peristalsis, a pattern of contraction in which a wave of contraction moves along the gut tube from one end to the other.
LINK
The structure and function of the vertebrate gut are detailed in Concept 30.4
In both cardiac muscle and smooth muscle, excitation–contraction coupling depends on Ca2+ influx into the sarcoplasm, as it does in skeletal muscle. Details of excitation–contraction coupling differ among the three types of vertebrate muscle, however.
Some insect flight muscle has evolved unique excitation–contraction coupling
Invertebrates have evolved a variety of specialized types of muscle. Here we discuss just three of them. Two of these, asynchronous flight muscle and catch muscle, are specialized to contract in unusual ways that are highly adaptive for the animals in which they occur. Another specialized type does not contract at all but instead has become modified to produce electricity!
The muscles that drive the wings during insect flight are of two types in terms of their excitation–contraction coupling. In about 25 percent of insect species, each contraction requires excitation. The sphinx moth in our opening photograph is one of these. Each excitation of a muscle cell brings about one contraction, meaning the ratio of excitations to contractions is 1:1. For muscle of this type to contract at a higher and higher frequency, all the processes of excitation–contraction coupling must take place faster and faster. At high frequencies, contraction becomes severely compromised in several ways as a result. Both muscular efficiency and power output diminish at high frequencies.
A different and unique form of muscle that has evolved only in insects solves these problems. It is termed asynchronous muscle because each excitation results in many contractions, meaning contractions are not synchronized in a 1:1 ratio with excitations. At high frequencies of contraction, asynchronous muscle maintains higher efficiency and greater power output than synchronous muscle. The reason asynchronous muscle has evolved in insects seems to be that it is well adapted in these ways to meeting the particular challenges of insect flight. Asynchronous muscle has evolved independently in insects about ten times and is found today in about 75 percent of insect species.
Catch muscle in clams and scallops stays contracted with little ATP use
A scallop or clam has a shell composed of two half-shells. One or two powerful adductor muscles connect these half-shells. Contraction of the adductor muscles pulls the two half-shells tightly together so that the animal becomes fully enclosed in its hard shell—a form of armor:

A scallop or clam’s adductor muscles sometimes need to remain contracted for long periods of time when predators are nearby.
The adductor muscles (and certain other muscles of mollusks) are able to enter a specialized state termed catch in which they maintain high contractile force for tens of minutes or even for hours with almost no use of ATP. The way catch works is not well understood. However, measurements show that catch muscles maintain contraction at a far lower ATP cost than most muscles. Catch is a highly controlled state that can be quickly ended by signals from the nervous system. Its energy-saving feature is adaptive for animals that have body armor that depends on muscular contraction to be effective.
Fish electric organs are composed of modified muscle
Electric eels (Electrophorus), which live in tropical rivers, can produce external voltage pulses as large as 700 volts. Electric rays (Torpedo) in the ocean also produce large external electric pulses, although they produce high electric currents (up to 20 amperes) rather than high voltages. Fish such as these use their electric pulses to stun prey. Many other fish produce small external electric pulses that they use to detect their surroundings for orientation.
The electric organs of nearly all electric fish have evolved from skeletal muscle and consist of modified muscle cells. These cells cannot contract because the contractile apparatus (e.g., actin and myosin) in each cell is poorly developed. However, the cell membranes of the muscle cells are unique in that the tiny voltage differences across the cell membranes of many cells all add together when the cells are excited during an electric pulse:
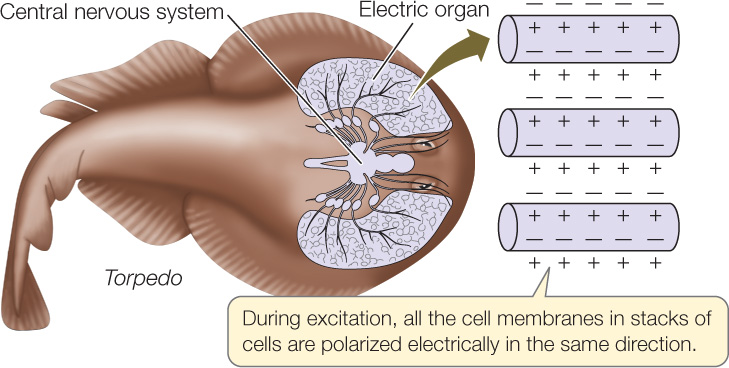
The voltage difference across each cell membrane is only about 0.1 V. In an electric organ, the voltage differences of many membranes add up, in the same way that the voltages of batteries add up when the batteries are connected in series. For this reason, an electric organ with 1,000 cell membranes can produce 100 V.
CHECKpointCONCEPT33.4
- Why does the heart keep beating in victims of neck fractures, although the victims stop breathing?
- What type of muscle would be used in each of the following activities: the extension of a lizard’s forelimbs during walking; the pumping of O2-poor blood from a person’s heart to his or her lungs; the expulsion of food during vomiting; the holding shut of a clam’s shell?
- If needed, could the modified muscle cells of fish electric organs be used to generate forces for swimming? Why or why not?
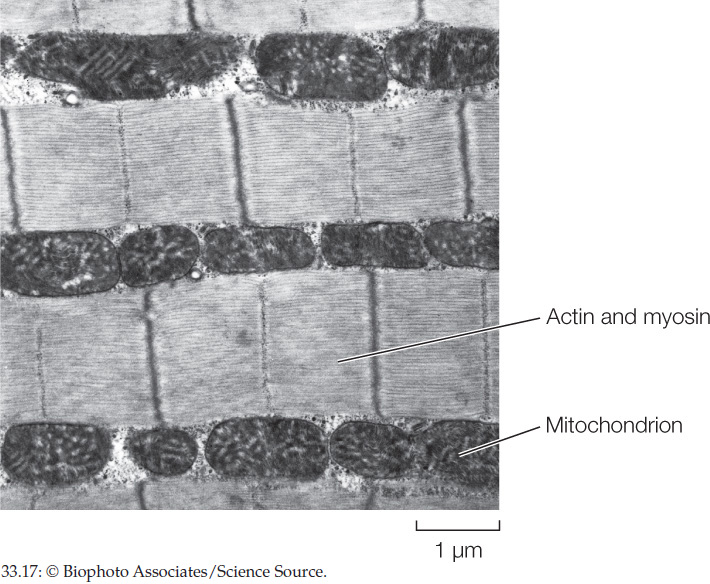
Why is it likely that available space inside cells has limited the contents of contractile proteins and mitochondria in high-performance muscles?
ANSWER Figure 33.17 is a highly magnified image of an insect flight muscle cell, obtained by electron microscopy. The inside of the cell is filled almost completely by mitochondria and contractile proteins. Open space is thus a scarce resource. Put simply, a high-performance muscle cell needs as large a set of contractile proteins as possible and as many mitochondria as possible—meaning there is a sort of “competition” for space in which the amounts of contractile proteins and mitochondria are each limited by space shortage. If, over evolutionary time, natural selection started to favor larger numbers of contractile protein molecules, the contractile proteins could edge out some of the mitochondria—jeopardizing the ability of the contractile proteins to get enough ATP. If natural selection started to favor more mitochondria, the mitochondria would edge out contractile proteins—jeopardizing the ability of the cell to use the ATP it could produce. The fact that space is limited has resulted in a sort of compromise in the use of space inside a high-performance muscle cell.