CONCEPT38.4 Gastrulation Sets the Stage for Organogenesis and Neurulation in Chordates
Organogenesis refers to the phase in development in which all of the organs and organ systems form. We will discuss the organs sequentially, but keep in mind that in an actual embryo, the processes of organogenesis occur simultaneously. Here we focus on organogenesis principally in the chordates, the large clade that includes the vertebrates and all other animals that possess a notochord. These animals also possess a dorsal nerve cord with a central cavity—a defining chordate characteristic (see Figure 16.4). The nerve cord and brain form by a process known as neurulation.
Each of the embryo’s three germ layers gives rise to a defined set of tissues and organs:
- Ectoderm, the outer germ layer, forms the epidermis of the skin and structures such as hair, claws, and sweat glands, the brain and all of the nervous system, and pigment cells that provide color to skin, hair, feathers, and more.
- Endoderm, the innermost germ layer, produces the lining of the digestive tract and the organs that arise from it, among them the lungs, liver, pancreas, and gall bladder.
- Mesoderm, the middle layer that surrounds the coelom, gives rise to the notochord, heart, blood and blood vessels, urogenital system, muscles, bones, and the dermis (the inner layer of the skin). In vertebrates, the dermis interacts with the overlying ectoderm (the epidermis) to produce many structures, including scales, teeth, hair, feathers, horns, sweat glands, and mammary glands.
Taking a big-picture view, the basic body plan can be seen as a tube within a tube. The outer tube, arising from ectoderm and mesoderm, is the body wall. The inner tube, arising from endoderm and mesoderm, is the gut. In between is the coelom. The major organs and organ systems develop as elaborations of this basic plan.
As development proceeds, cells become recognizable as belonging to a particular type (neurons, blood cells, liver cells and so on). When a cell displays its final specialized characteristics, it is said to be differentiated. Differentiation takes place gradually throughout development and often into adult life. For example, mammalian red blood cells contain hemoglobin and lack nuclei. Old red blood cells are replaced continuously throughout the animal’s life as new cells are produced and differentiate.
The notochord induces formation of the neural tube
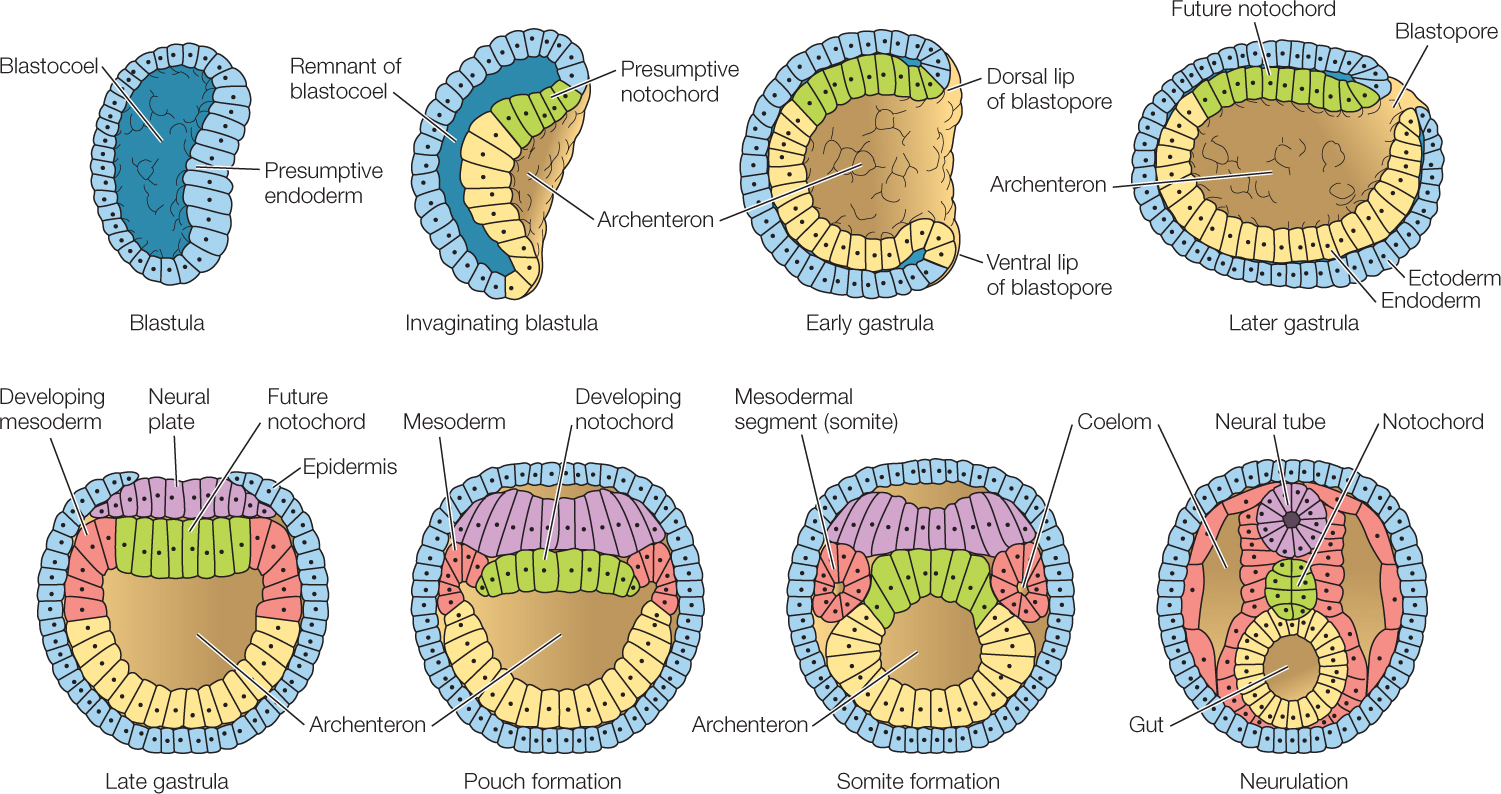
In many chordates, some of the cells that move through the blastopore during gastrulation separate from the roof of the archenteron (FIGURE 38.13) and form the notochord (see Figure 23.35). In amphibians, for example, these are cells from the dorsal blastopore lip. In addition to providing structural support, the notochord plays a critical role during development because it induces the overlying ectoderm to form the dorsal nerve cord. Induction refers to the ability of one tissue to direct or lead another along a specific pathway of development. We now know that this is done by chemical signaling. Induction is one of the most important mechanisms in biology. It probably takes place in all animals and plays a role in the formation of most organ systems.
LINK
Some of the molecular mechanisms of induction are discussed in Concept 14.2; see especially Figure 14.9
Under the influence of the developing notochord, a portion of the dorsal ectoderm flattens and begins to roll up, forming first a neural plate and then a neural groove. When the edges of the groove grow together, they fuse to form the neural tube, and ectoderm closes over them. At its anterior end, the vertebrate neural tube forms three swellings, which will become the major divisions of the adult brain: the hindbrain, midbrain, and forebrain. The rest of the neural tube will become the spinal cord. If the neural tube does not close properly, serious problems result. Examples in humans include spina bifida (an opening in the spine that allows the spinal cord to protrude) and anencephaly (absence of a major portion of the brain).
Gastrulation sets the stage for organogenesis and neurulation in chordates
What about the amphibian dorsal blastopore lip makes it so crucial to “organizing” the three germ layers? The distribution of β-catenin in the late blastula corresponds to the location of this organizer in the early gastrula. A series of experiments endeavored to ascertain whether β-catenin is “necessary and sufficient” for subsequent functions of the organizer. Review the results of these experiments in the table and answer the questions.
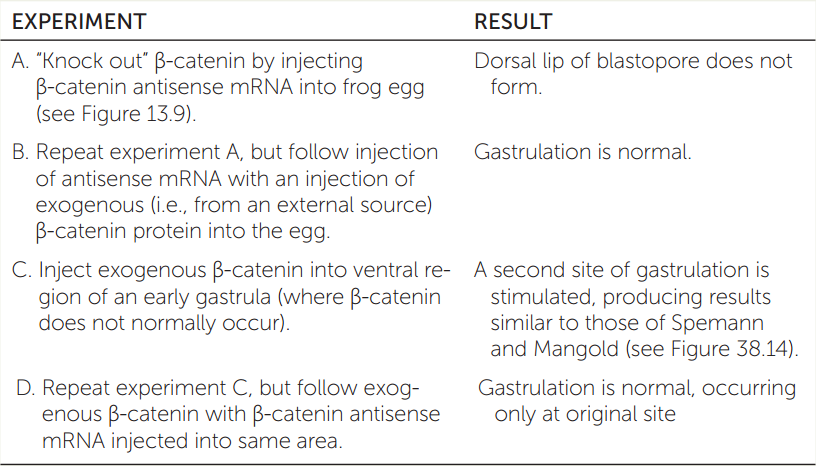
- Which experiment shows that β-catenin is necessary for induction of the organizer? Explain.
- Which experiment shows that β-catenin is sufficient (enough) for induction of the organizer? Explain.
- What was the purpose of experiment B? Of experiment D?
- This series of experiments demonstrates what is sometimes called the “find it, lose it, move it” approach to understanding a gene’s function. Explain why this catchy phrase is appropriate.
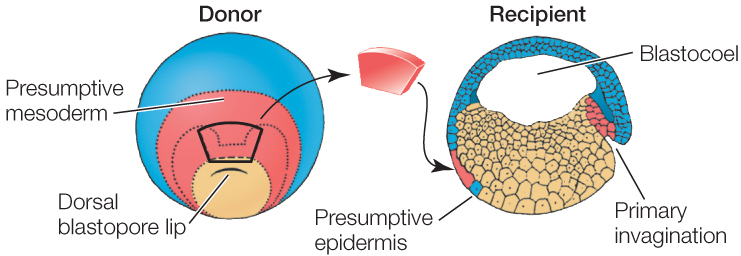
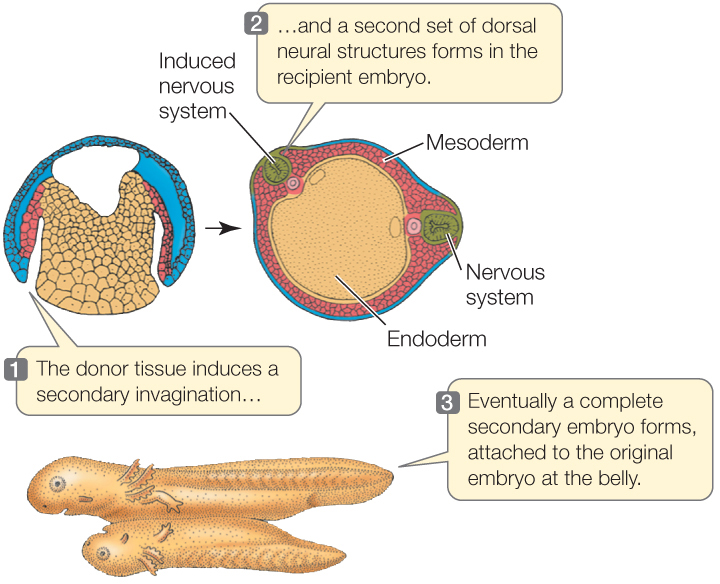
Much of our understanding of induction comes from work that began on amphibian embryos in the early twentieth century and has continued since. When Hans Spemann and his student Hilde Mangold were studying the dorsal lip of the blastopore in frog eggs, they transplanted this region to another embryo at the same stage of development. The results were momentous. This small piece of tissue stimulated a second site of gastrulation, and a second complete embryo formed (FIGURE 38.14). Because the dorsal blastopore lip of amphibians was apparently capable of inducing host tissue to form an entire embryo, Spemann and Mangold dubbed it the organizer, and its action became known as primary embryonic induction. We now know that the dorsal blastopore lip is not responsible for the first inductive event in development, but because of its extreme importance, it is still referred to as the organizer. For more than 80 years, the organizer has been an active area of research. Hensen’s node, which we discussed in Concept 38.3, acts as an organizer in birds, and a homologous region does so in mammals.
Investigation
HYPOTHESIS
Cytoplasmic factors in the early dorsal blastopore lip organize cell differentiation in amphibian embryos.
- Excise a patch of tissue from above the dorsal blastopore lip of an early gastrula stage salamander embryo (the donor).
- Transplant the donor tissue onto a recipient embryo at the same stage. The donor tissue is transplanted onto a region of ectoderm that should become epidermis (skin).
CONCLUSION
The cells of the dorsal blastopore lip can induce other cells to change their developmental fates.
aH. Spemann and H. Mangold. 1924. Roux’ Arch. Entw. Mech. 100: 599–638. Viktor Hamburger’s translation appeared in Foundations of Experimental Embryology, 1964, (B.H. Willier and J.M. Oppenheimer, eds.), pp. 146–184.
Molecular biologists are making great strides in working out the molecular mechanisms of the organizer. Thanks to a series of experiments in the 1980s and 1990s, we now know that the protein β-catenin plays a key role in determining which cells become the organizer. It does this by triggering a complex series of interactions between various transcription factors and growth factors that control gene expression. Several of these transcription factors have been identified.
During the last century, clever experiments revealed the source and approximate molecular size of many inductive factors. Now, modern techniques are enabling scientists to determine the identity of many such molecules. One of the best-studied examples is the induction of the dorsal nerve cord in amphibians.
The development of the central nervous system depends on the activation of a specific genetic cascade within certain ectodermal cells that will become neuronal instead of epidermal. Work on amphibian embryos has shown that specific polypeptides released from the developing notochord act as inducers that control this activation. Two of these polypeptides that induce the formation of the neural tube are appropriately named Noggin and Chordin. They act, not by direct activation of genes, but rather by inhibiting the activity of another substance, the TGF-β bone morphogenetic protein BMP4 (FIGURE 38.15). Among its many actions, BMP4 induces the ectoderm to form epidermis. In the absence of BMP4, ectoderm forms neural tissue. By blocking the effect of BMP4, Chordin and Noggin allow the ectoderm to carry out its “default” differentiation of becoming neural.
Investigation
HYPOTHESIS
The default state of amphibian dorsal ectoderm is neural; it is induced by underlying mesoderm to become epidermis.
- Excise animal caps (presumptive ectoderm) from amphibian blastulas. Culture presumptive mesodermal cells from early gastrulas and extract BMP4. BMP4 is a growth factor released from the notochord that induces overlying ectoderm to become skin.
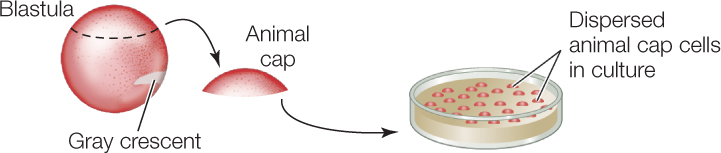
- Prepare four separate cultures of embryonic ectodermal cells. Incubate with no additions (control); with BMP4 from step 1; with a BMP4 inhibitor; and with both molecules.
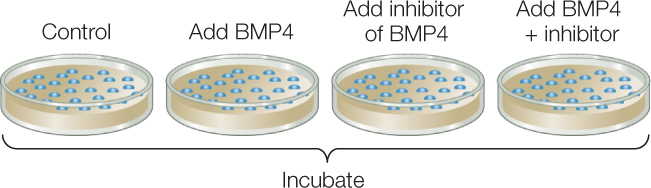
- After incubation, extract mRNAs from the cultured ectodermal cells and run on gels to reveal expression of NCAM (a neural tissue marker) and keratin (an epidermal tissue marker).
Gels reveal that different treatments with BMP4 and its inhibitor alter patterns of gene expression in cultured ectodermal cells.
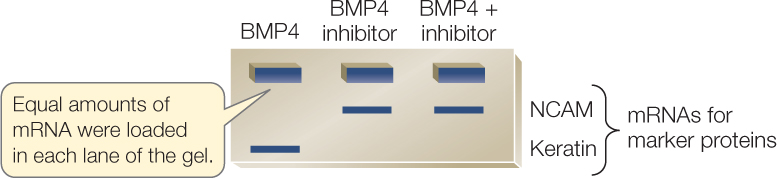
CONCLUSION
The organizer cells secrete an inhibitor of BMP4.
ANALYZE THE DATA
Use the gel results shown above to answer the questions.
- Does BMP4 induce expression of neural-specific or epidermal-specific genes?
- Does BMP4 block any gene expression in the ectodermal cells?
- What is the evidence that BMP4 has an inductive and/or an inhibitory effect on gene expression in ectodermal cells?
- Do these results support the hypothesis?
aP. A. Wilson and A. Hemmati-Brivanlou. 1995. Nature 376: 331–333.
Another important factor controlling differentiation of the neural tube is Sonic hedgehog, symbolized Shh, the transcription factor mentioned at the start of this chapter. Shh is released by the notochord and diffuses into the ventral region of the neural tube, where it directs the development of this region into the ventral structures and circuits of the spinal cord. These circuits will carry the major output from the motor pathways of the nervous system.
LINK
Sonic hedgehog also plays an important role in the development of vertebrate limbs; see Concept 14.3
As the neural tube is closing, certain cells dissociate from it and come to lie between the neural tube and the overlying epidermis. These are known as neural crest cells (FIGURE 38.16). They are star-shaped cells that are termed “pluripotent” because they can differentiate in many different ways. Neural crest cells migrate extensively and have amazing developmental capacities. Not only do they form sensory neurons and major parts of the autonomic nervous system, but they are also responsible for most of the skull bones, the pigment cells, and many other structures. Without neural crest cells, vertebrates would not have heads, as we recognize them.
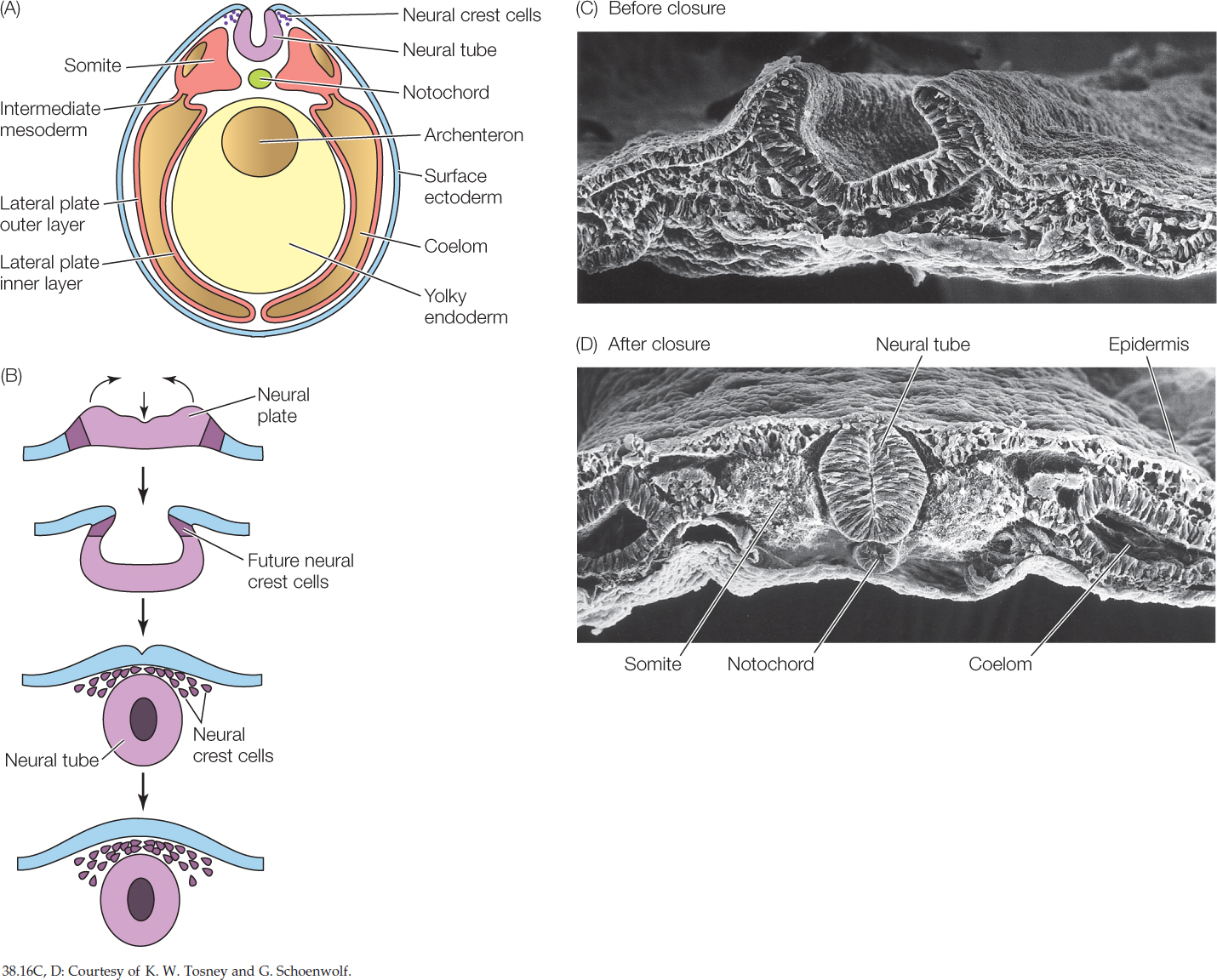
Mesoderm forms tissues of the middle layer
On either side of the forming notochord, three regions of mesoderm can be recognized—somites, intermediate mesoderm, and lateral plate mesoderm (see Figure 38.16). The vertebrate body plan, like that of annelids and arthropods, consists of repeating segments that are modified during development. These segments are first seen as the blocks of cells known as somites (FIGURE 38.17; also see Figure 38.16), then later as the repeating patterns of vertebrae, ribs, nerves, and muscles along the anterior–posterior axis as differentiation proceeds.
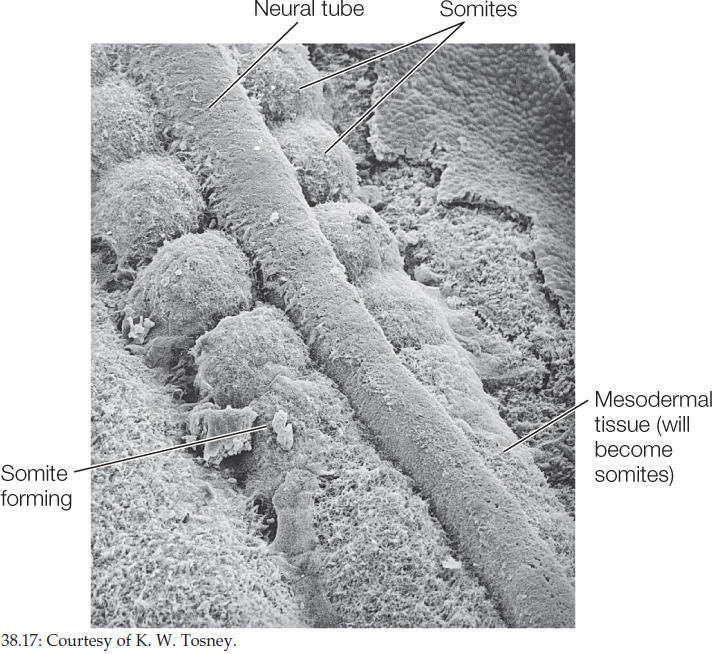
Somites form bones, cartilage, skeletal muscle, and the dermis of the skin.
Intermediate mesoderm forms much of our urinary and reproductive system.
Lateral plate mesoderm surrounds the coelom and lines the vertebrate body cavity (the peritoneal cavity). The peritoneal cavity makes up much of the coelom. The thin tissue layer that surrounds the peritoneal cavity is the peritoneum. (You may have heard of peritonitis, a severe inflammation of the covering of the internal organs. A major cause is the entry of bacteria into the peritoneal cavity, as occurs when an appendix ruptures.) The inner layer of lateral plate mesoderm lies close to the endoderm. It forms part of the peritoneum, the muscles of the digestive tract, and most of the circulatory system, including the heart. The outer layer of lateral plate mesoderm lies adjacent to the ectoderm and forms part of the peritoneum and some of the muscle of the body wall (see Figure 38.16).
CHECKpointCONCEPT38.4
- What two major mechanisms are responsible for determining the differentiated state of a cell?
- What is the role of the notochord in neurulation?
- A chordate synapomorphy (a shared derived trait) is a hollow dorsal nerve chord (see Concept 23.5). How does the process of neurulation explain why the spinal cord has a central cavity?
- What major tissues does ectoderm form? What three major mesodermal regions form in chordates, and what does each form?
We have seen how the three germ layers form the organs and tissues of the vertebrate embryo. Next we will focus on how they also protect and nourish the embryo in many of the animals with which you are most familiar.