8.1 RNA
Early investigators had good reason for thinking that information is not transferred directly from DNA to protein. In a eukaryotic cell, DNA is found in the nucleus, whereas protein is synthesized in the cytoplasm. An intermediate is needed.
Early experiments suggest an RNA intermediate
In 1957, Elliot Volkin and Lawrence Astrachan made a significant observation. They found that one of the most striking molecular changes that takes place when E. coli is infected with the phage T2 is a rapid burst of RNA synthesis. Furthermore, this phage-
Volkin and Astrachan demonstrated the rapid turnover of RNA by using a protocol called a pulse-chase experiment. To conduct a pulse–
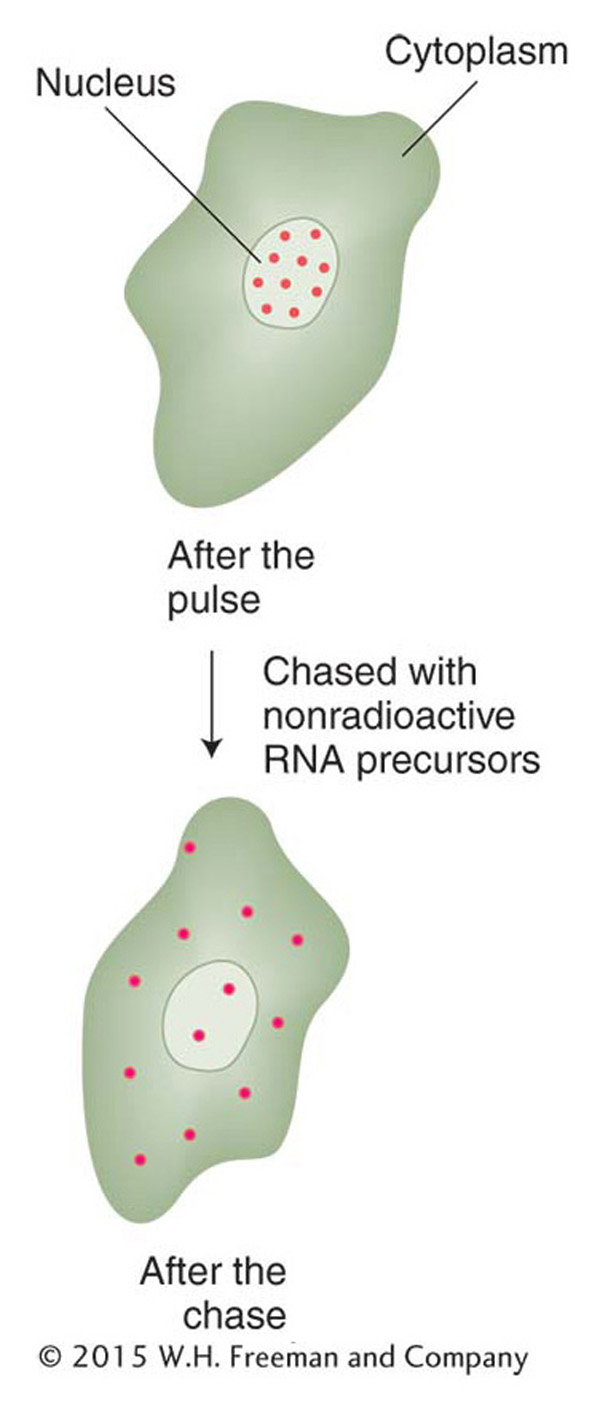
A similar experiment can be done with eukaryotic cells. Cells are first pulsed with radioactive uracil and, after a short time, they are transferred to medium with unlabeled uracil. In samples taken after the pulse, most of the label is in the nucleus. In samples taken after the chase, the labeled RNA is also found in the cytoplasm (Figure 8-1). Apparently, in eukaryotes, the RNA is synthesized in the nucleus and then moves into the cytoplasm, where proteins are synthesized. Thus, RNA is a good candidate for an information-
Properties of RNA
Let’s consider the general features of RNA. Although both RNA and DNA are nucleic acids, RNA differs from DNA in several important ways:
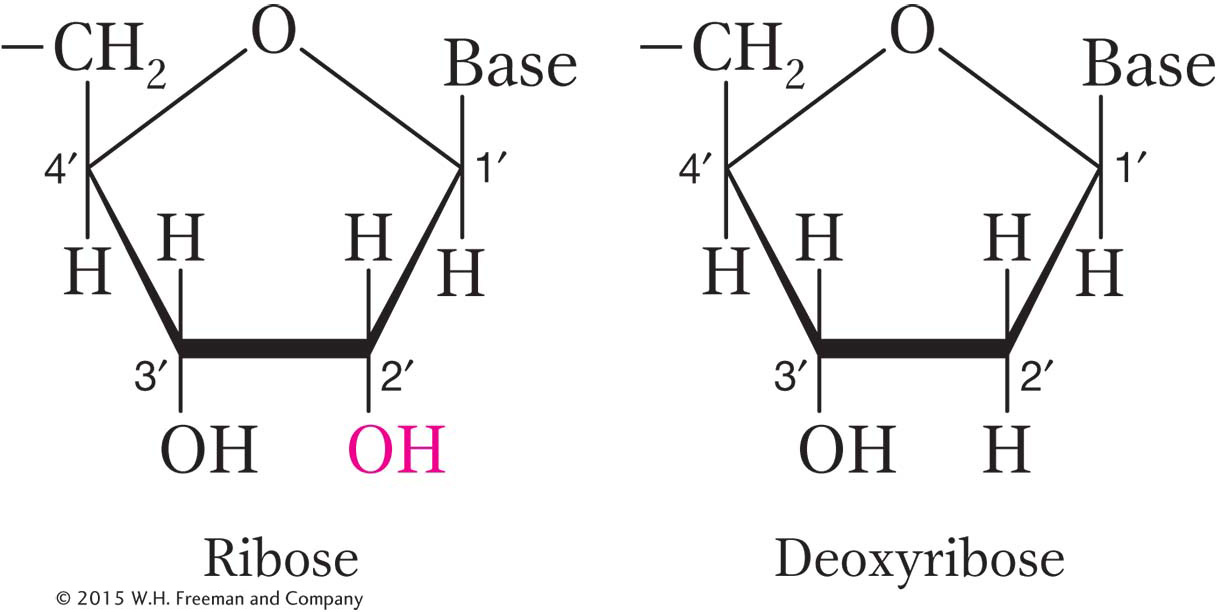
RNA has ribose sugar in its nucleotides, rather than the deoxyribose found in DNA. As the names suggest, the two sugars differ in the presence or absence of just one oxygen atom. The RNA sugar contains a hydroxyl group (OH) bound to the 2′-carbon atom, whereas the DNA sugar has only a hydrogen atom bound to the 2′-carbon atom.
RNA is usually a single-
stranded nucleotide chain, not a double helix like DNA. A consequence is that RNA is more flexible and can form a much greater variety of complex three- dimensional molecular shapes than can double- stranded DNA. An RNA strand can bend in such a way that some of its own bases pair with each other. Such intramolecular base pairing is an important determinant of RNA shape. As you will see later in this chapter, the presence of the hydroxyl group at the 2′-carbon atom facilitates the action of RNA in many important cellular processes.
Like an individual DNA strand, a strand of RNA is formed of a sugar–
phosphate backbone, with a base covalently linked at the 1′ position on each ribose. The sugar– phosphate linkages are made at the 5′ and 3′ positions of the sugar, just as in DNA; so an RNA chain will have a 5′ end and a 3′ end. 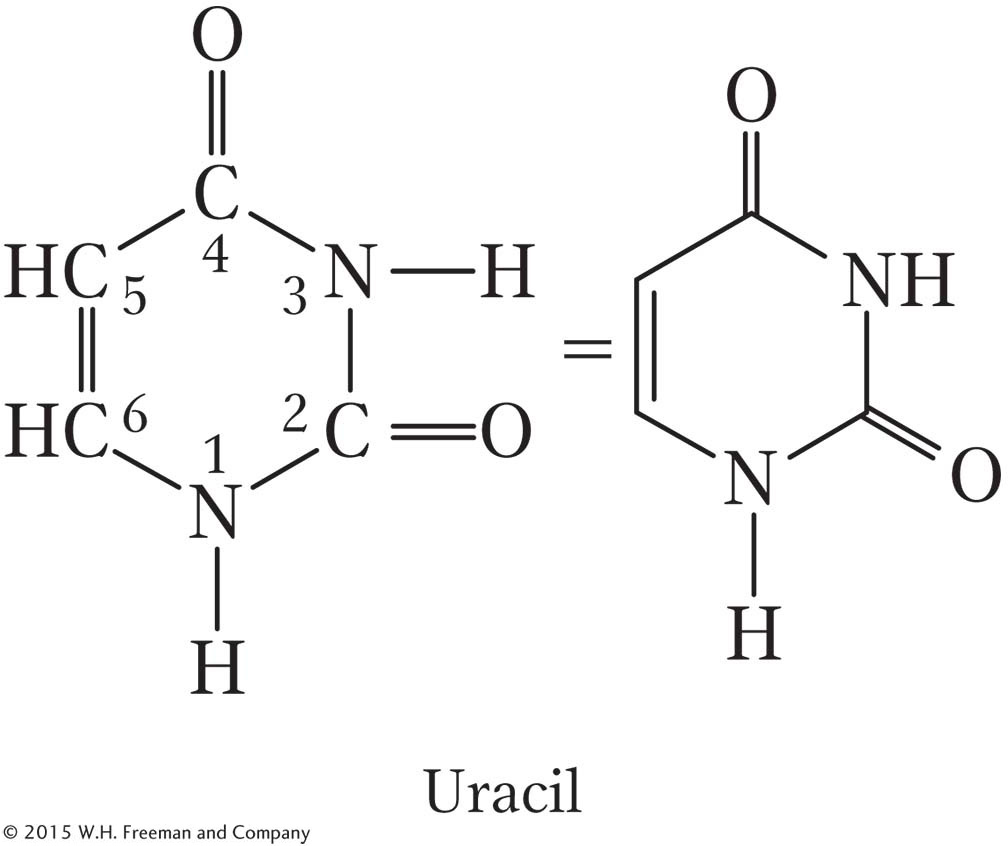
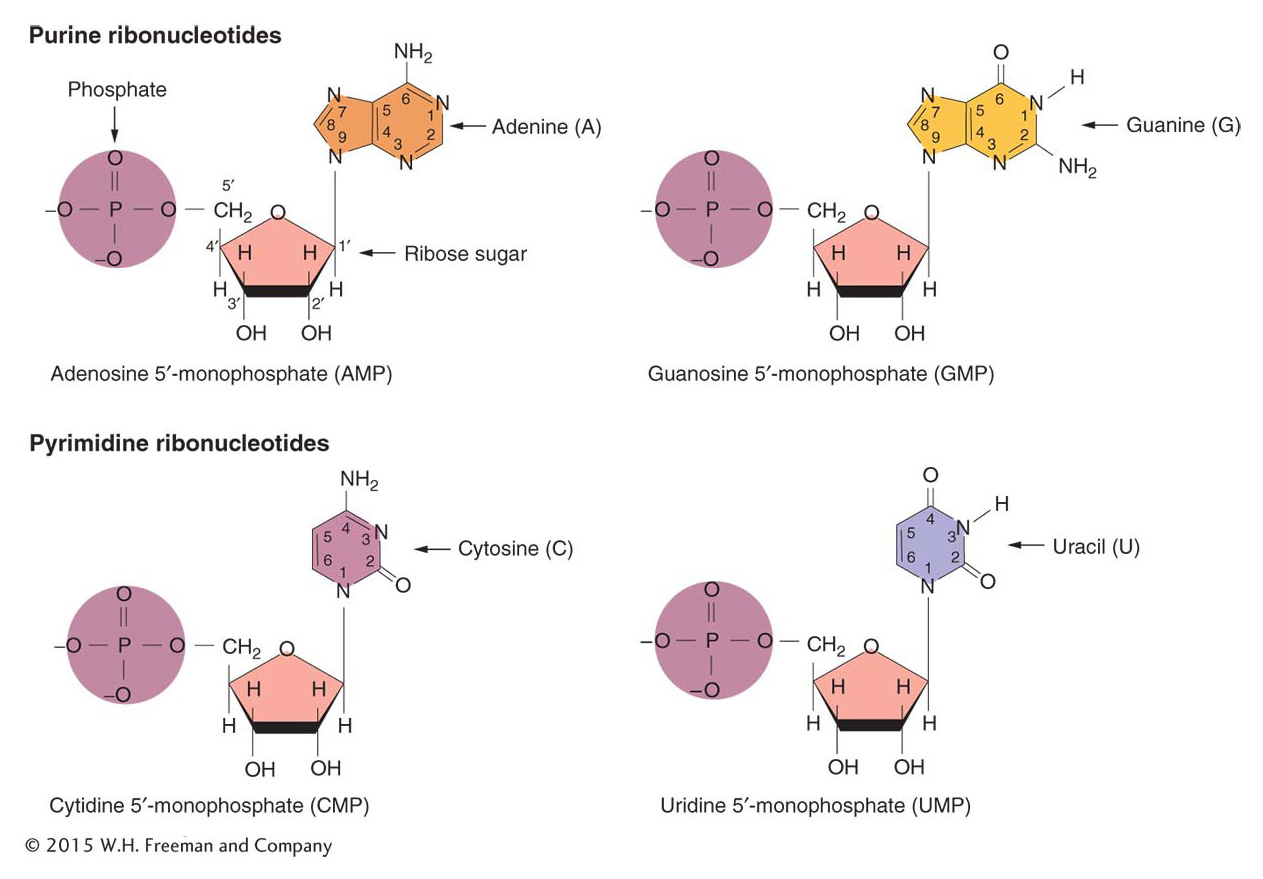
RNA nucleotides (called ribonucleotides) contain the bases adenine, guanine, and cytosine, but the pyrimidine base uracil (abbreviated U) is present instead of thymine.
Uracil forms two hydrogen bonds with adenine just as thymine does. Figure 8-2 shows the four ribonucleotides found in RNA.
In addition, uracil is capable of base pairing with G. The bases U and G form base pairs only during RNA folding and not during transcription. The two hydrogen bonds that can form between U and G are weaker than the two that form between U and A. The ability of U to pair with both A and G is a major reason why RNA can form extensive and complicated structures, many of which are important in biological processes.
RNA—
like protein, but unlike DNA— can catalyze biological reactions. The name ribozyme was coined for the RNA molecules that function like protein enzymes.
Classes of RNA
RNAs can be grouped into two general classes. One class of RNA encodes the information necessary to make polypeptide chains (proteins). We refer to this class as messenger RNA (mRNA) because, like a messenger, these RNAs serve as the intermediary that passes information from DNA to protein. We refer to the other class as functional RNA because the RNA does not encode information to make protein. Instead, the RNA itself is the final functional product.
Messenger RNA The steps through which a gene influences phenotype are called gene expression. For the vast majority of genes, the RNA transcript is only an intermediate necessary for the synthesis of a protein, which is the ultimate functional product that influences phenotype.
Functional RNA As more is learned about the intimate details of gene expression and regulation, it becomes apparent that functional RNAs fall into a variety of classes and play diverse roles. Again, it is important to emphasize that functional RNAs are active as RNA; they are never translated into polypeptides.
The main classes of functional RNAs contribute to various steps in the transfer of information from DNA to protein, in the processing of other RNAs, and in the regulation of RNA and protein levels in the cell. Two such classes of functional RNAs are found in both prokaryotes and eukaryotes: transfer RNAs and ribosomal RNAs.
Transfer RNA (tRNA) molecules are responsible for bringing the correct amino acid to the mRNA in the process of translation.
Ribosomal RNA (rRNA) molecules are the major components of ribosomes, which are large macromolecular machines that guide the assembly of the amino acid chain by the mRNAs and tRNAs.
The entire collection of tRNAs and rRNAs are encoded by a small number of genes (a few tens to a few hundred at most). However, though the genes that encode them are few in number, rRNAs account for a very large percentage of the RNA in the cell because they are both stable and transcribed into many copies.
Another class of functional RNAs participate in the processing of RNA and are specific to eukaryotes:
Small nuclear RNAs (snRNAs) are part of a system that further processes RNA transcripts in eukaryotic cells. Some snRNAs unite with several protein subunits to form the ribonucleoprotein processing complex (the spliceosome) that removes introns from eukaryotic mRNAs.
Finally, a large group of functional RNAs suppress the expression of genes at many levels and also maintain genome stability. Three classes of these functional RNAs may be encoded by large parts of eukaryotic genomes: microRNAs, small interfering RNAs, and piwi-
MicroRNAs (miRNAs) have recently been recognized to have a widespread role in regulating the amount of protein produced by many eukaryotic genes.
Small interfering RNAs (siRNAs) and piwi-interacting RNAs (piRNAs) help protect the integrity of plant and animal genomes. siRNAs inhibit the production of viruses, while both siRNAs and piRNAs prevent the spread of transposable elements to other chromosomal loci. siRNAs restrain transposable elements in plants, and piRNAs perform the same function in animals.
Long noncoding RNAs (lncRNAs), or sometimes just abbreviated ncRNAs were recently found to be transcribed from most regions of the genomes of humans and other animals and plants. While a few lncRNAs play a role in classic genetic phenomena such as dosage compensation (see Chapter 12), the function, if any, of most lncRNAs is currently unknown.
Because protein synthesis and mRNA processing occur throughout the lifetime of most cells, tRNA, rRNA, and snRNAs are always needed. As such, these RNAs are continuously synthesized (their transcription is said to be constitutive). In contrast, miRNAs, siRNAs, piRNAs, and lncRNAs are transcribed and/or processed from larger transcripts intermittently, only when they are needed to fulfill their roles in protecting the genome and regulating gene expression.