Chapter Introduction
Proteins and Their
Synthesis
CHAPTER
9
LEARNING OUTCOMES
After completing this chapter, you will be able to
Compare the sequences of a gene and its protein to evaluate their relationship.
Examine and explain the experimental findings that the genetic code is nonoverlapping and degenerate.
Understand that although translation is conserved across all of life, there are some important differences between prokaryotes and eukaryotes.
Compare and contrast the critical roles played by two functional RNAs—
ribosomal RNA and tRNA— in protein synthesis. Provide evidence that the ribosomal RNA, not the ribosomal proteins, carries out the key steps in translation.
Compare different types of posttranslational processing and their importance for protein function.
In an address to Congress in 1969, William Stewart, Surgeon General of the United States, said, “It is time to close the book on infectious diseases. The war against pestilence is over.” At the time, his claim of victory was not an unreasonable boast. In the preceding two decades, three infectious diseases that had plagued humankind for centuries—
Unfortunately, William Stewart’s claim of victory in the battle against infectious disease was premature. The overuse of antibiotics worldwide has spurred the evolution of resistant bacterial strains. For example, each year, more than 2 million hospital patients in the United States acquire an infection that is resistant to antibiotics and 90,000 die as a result. How did resistance develop so quickly? Will infectious disease be, once again, a significant cause of human mortality? Or will scientists be able to use their understanding of resistance mechanisms to develop more durable antibiotics?
To answer these questions, scientists have focused on the cellular machinery that is targeted by antibiotics. More than half of all antibiotics currently in use target the bacterial ribosome, the site of protein synthesis in prokaryotes. In this chapter, you will learn that scientists have had incredible success with the use of a technique called X-
Chapters 7 and 8 described how DNA is copied from generation to generation and how RNA is synthesized from specific regions of DNA. We can think of these processes as two stages of information transfer: replication (the synthesis of DNA) and transcription (the synthesis of an RNA copy of a part of the DNA). In this chapter, you will learn about the final stage of information transfer: translation (the synthesis of a polypeptide directed by the RNA sequence).
As you learned in Chapter 8, RNA transcribed from genes is classified as either messenger RNA (mRNA) or functional RNA. In this chapter, we will see the fate of both RNA classes. The vast majority of genes encode mRNAs whose function is to serve as an intermediate in the synthesis of the ultimate gene product, protein. In contrast, recall that functional RNAs are active as RNAs; they are never translated into proteins. The main classes of functional RNAs are important actors in protein synthesis. They include transfer RNAs and ribosomal RNAs.
Transfer RNA (tRNA) molecules are the adapters that translate the threenucleotide codon in the mRNA into the corresponding amino acid, which is brought by the tRNA to the ribosome in the process of translation. The tRNAs are general components of the translation machinery; a tRNA molecule can bring an amino acid to the ribosome for the purpose of translating any mRNA.
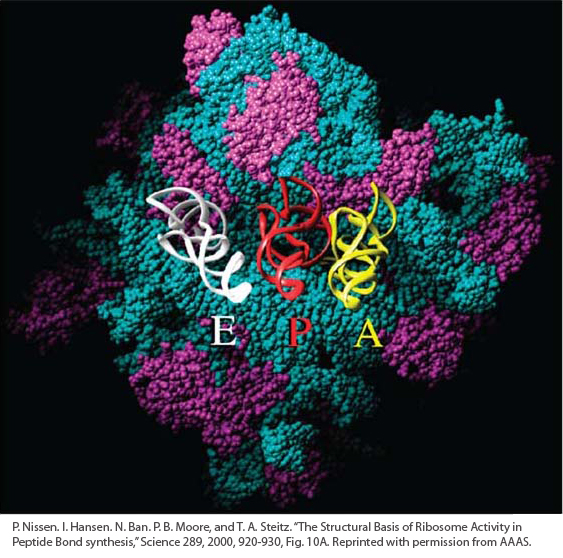
Page 321Ribosomal RNAs (rRNAs) are the major components of ribosomes, which are large macromolecular complexes that assemble amino acids to form the protein whose sequence is encoded in a specific mRNA. Ribosomes are composed of several types of rRNA and scores of different proteins. Like tRNA, ribosomes are general in function in the sense that they can be used to translate the mRNAs of any protein-
coding gene.
Although most genes encode mRNAs, functional RNAs make up, by far, the largest fraction of total cellular RNA. In a typical actively dividing eukaryotic cell, rRNA and tRNA account for almost 95 percent of the total RNA, whereas mRNA accounts for only about 5 percent. Two factors explain the abundance of rRNAs and tRNAs. First, they are much more stable than mRNAs, and so these molecules remain intact much longer. Second, because an actively dividing eukaryotic cell has tens of thousands of ribosomes, the transcription of rRNA and tRNA genes constitutes more than half of the total nuclear transcription in active eukaryotic cells and almost 80 percent of transcription in yeast cells.
The components of the translational machinery and the process of translation are very similar in prokaryotes and eukaryotes. The major feature that distinguishes translation in prokaryotes from that in eukaryotes is the location where transcription and translation take place in the cell: the two processes take place in the same compartment in prokaryotes, whereas they are physically separated in eukaryotes by the nuclear membrane. After extensive processing, eukaryotic mRNAs are exported from the nucleus for translation on ribosomes that reside in the cytoplasm. In contrast, transcription and translation are coupled in prokaryotes: translation of an RNA begins at its 5′ end while the rest of the mRNA is still being transcribed.