10-5 Auditory Communication in Nonhuman Species
Sound has survival value. You will appreciate this if you’ve ever narrowly escaped becoming an accident statistic by crossing a busy intersection on foot while listening to a music player or talking on a cell phone. Audition is as important a sense to many animals as vision is to humans. Many animals also communicate with other members of their species by using sound, as humans do.
Here we consider just two types of auditory communication in nonhumans: birdsong and echolocation. Each provides a model for understanding different aspects of brain–
Birdsong
Of about 8500 living bird species, about half are considered songbirds. Birdsong has many functions, including attracting mates (usually employed by males), demarcating territories, and announcing location or even just presence. Although all birds of the same species have a similar song, the song’s details vary markedly from region to region, much as dialects of the same human language vary.
Parallels Between Birdsong and Language
Figure 10-23 shows sound wave spectrograms for the songs of male white-
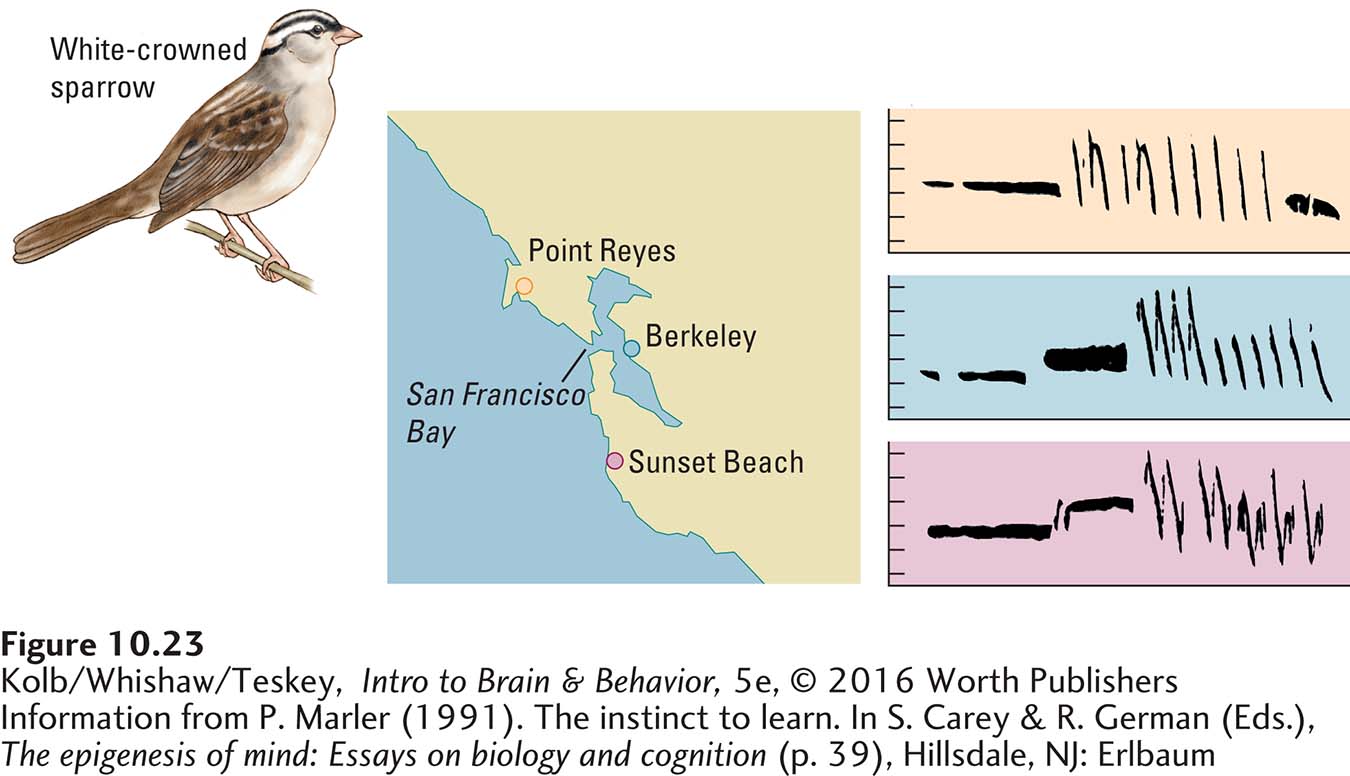
These gene–
Birdsong and human language have broad similarities beyond regional variation. Both appear to be innate yet are sculpted by experience. Both are diverse and can vary in complexity. Humans seem to have a basic template for language that is programmed into the brain, and experience adds a variety of specific structural forms to this template.
If a young bird is not exposed to song until it is a juvenile and then listens to recordings of birdsongs of various species, the young bird shows a general preference for its own species’ song. This preference must mean that each bird has a species-
Another broad similarity between birdsong and human language is their great diversity. Among birds, diversity is apparent in the sheer number of songs that a species possesses. Species such as the white-
The number of syllables in birdsong also varies greatly, ranging from 30 for the canary to about 2000 for the brown thrasher. Similarly, even though all modern human languages are equally complex, they vary significantly in the type and number of elements they employ. The number of meaningful patterns in speech sounds in human languages ranges from about 15 for some Polynesian languages to about 100 for some dialects spoken in the Caucasus Mountains. English has 24.
A final broad similarity between birdsong and human language lies in how they develop. In many bird species, song development is heavily influenced by experience during a critical period, just as language development is in humans. Birds also go through stages in song development, just as humans go through stages in language development. Hatchlings make noises that attract their parents’ attention, usually for feeding, and human babies emit cries to signal hunger, among other things.
The fledgling begins to make noises that Charles Darwin compared to the prespeech babbling of human infants. These noises, called subsong, are variable in structure, low in volume, and often produced as the bird appears to doze. Presumably, subsong, like human babbling, is practice for the later development of adult communication after the bird has left the nest.
As a young bird matures, it starts to produce sound wave patterns that contain recognizable bits of the adult song. Finally, the adult song emerges. In most species, the adult song remains remarkably stable, although a few species, such as canaries, can develop a new song every year that replaces the previous year’s song.
Neurobiology of Birdsong
The neurobiology of birdsong has been a topic of intense research, partly because it provides an excellent model of brain changes that accompany learning and partly because it offers insight into how sex hormones influence behavior. In the 1970s, Fernando Nottebohm and his colleagues first identified the major structures controlling birdsong, illustrated in Figure 10-24 (Nottebohm & Arnold, 1976). The largest structures are the higher vocal control center (HVC) and the nucleus robustus archistriatalis (RA). The axons of the HVC connect to the RA, which in turn sends axons to the 12th cranial nerve. This nerve controls the muscles of the syrinx, the structure that actually produces the song.
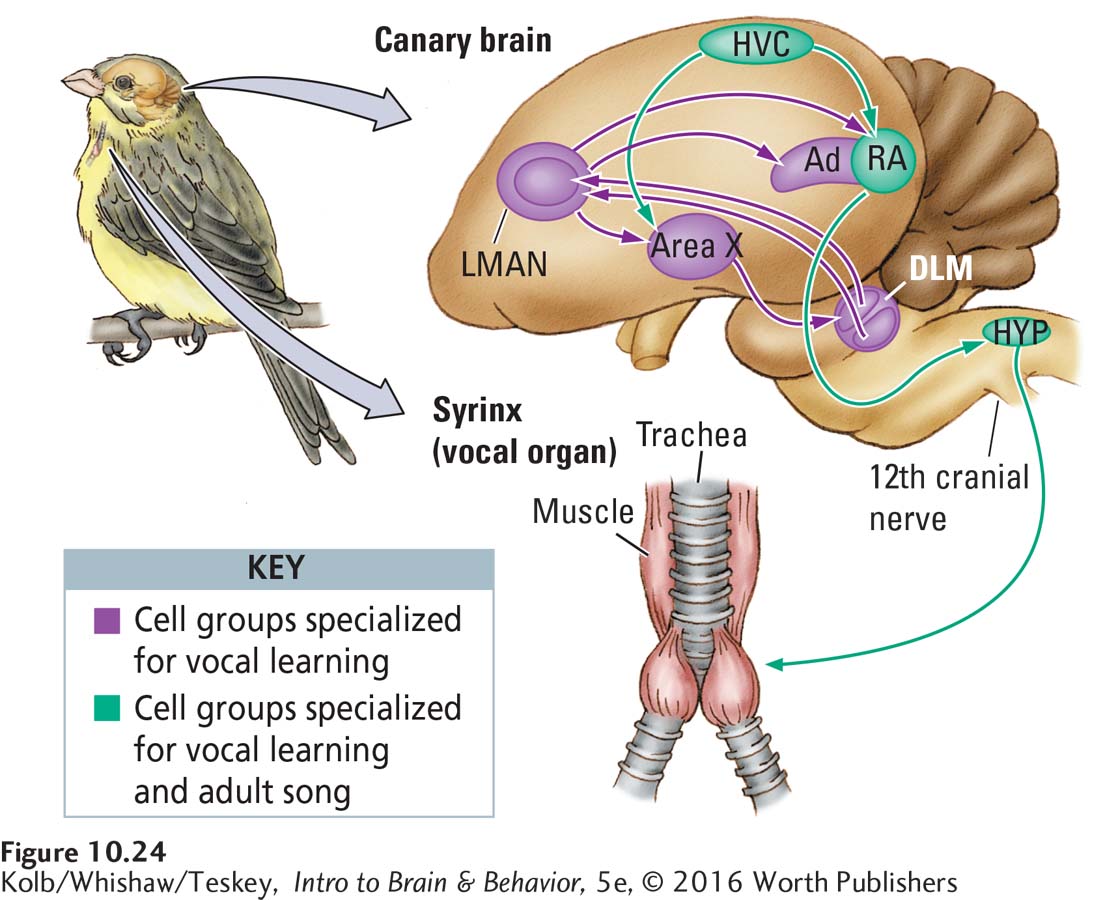
The HVC and RA have several important and some familiar characteristics:
Shown in Figure 8-30, a rare gyandromorph zebra finch exhibits physical characteristics of both sexes.
The structures are asymmetrical in some bird species, with those in the left hemisphere larger than those on the right. In many cases, this asymmetry is similar to the lateralized control of language in humans: if the left-
hemisphere pathways are damaged, the bird stops singing, but similar injury in the right hemisphere has no effect on song. Birdsong structures are sexually dimorphic, that is, much larger in males than in females. In male canaries, the structures are five times as large as in the female. This sex difference is due to the hormone testosterone in males. Injection of testosterone into female birds causes the song-
controlling nuclei to increase in size. The size of the birdsong-
controlling nuclei is related to singing skill. Unusually talented singers among male canaries tend to have larger HVCs and RAs than do less- gifted singers. Page 351The HVC and RA contain cells that produce birdsong as well as cells responsive to hearing song, especially the song of a bird’s own species.
The same structures therefore play a role in both song production and song perception. This avian neural anatomy is comparable to the overlapping roles of Broca’s and Wernicke’s areas in language perception and production in humans.
Echolocation in Bats
Dolphins use an auditory strategy similar to bats, but in water. Focus 10-2 profiles human echolocators.
Next to rodents, bats are the most numerous mammalian order. The two general groups, or suborders, are the smaller bats (Microchiroptera) and the larger fruit-

Most of the 680 species of Microchiroptera feed on insects. Others live on blood (vampire bats), and some catch frogs, lizards, fishes, birds, and small mammals. These bats’ auditory system is highly specialized to use echolocation not only to locate targets in the dark but also to analyze the targets’ features as well as environmental features in general. Through echolocation, these bats identify prey, navigate through the leaves of trees, and locate suitable landing surfaces. Echolocation in the Microchiroptera works rather like sonar. The bat larynx emits bursts of sound waves at ultrasonic frequencies. The waves bounce off objects and return to the bat’s ears, allowing the animal to identify what is in the surrounding environment. The bat, in other words, navigates by the echoes it hears, differentiating among the various characteristics of the echoes.
Moving objects (such as insects) give off a moving echo, smooth objects a different echo from rough objects, and so on. A key component of the bats’ echolocation system is analysis of differences in echo return times. Close objects return echoes sooner than more distant objects do, and the textures of various objects’ surfaces impose minute differences in return times.
A bat’s cries are short (ranging from 0.3 to 200 milliseconds) and high frequency (12,000 to 200,000 Hz, charted in Figure 10-4). Most of this range lies at too high a frequency for the human ear to detect. Different bat species produce sound waves of different frequencies that depend on the animal’s ecology. Bats that catch prey in the open use different frequencies from those used by bats that catch insects in foliage and from those used by bats that hunt prey on the ground.
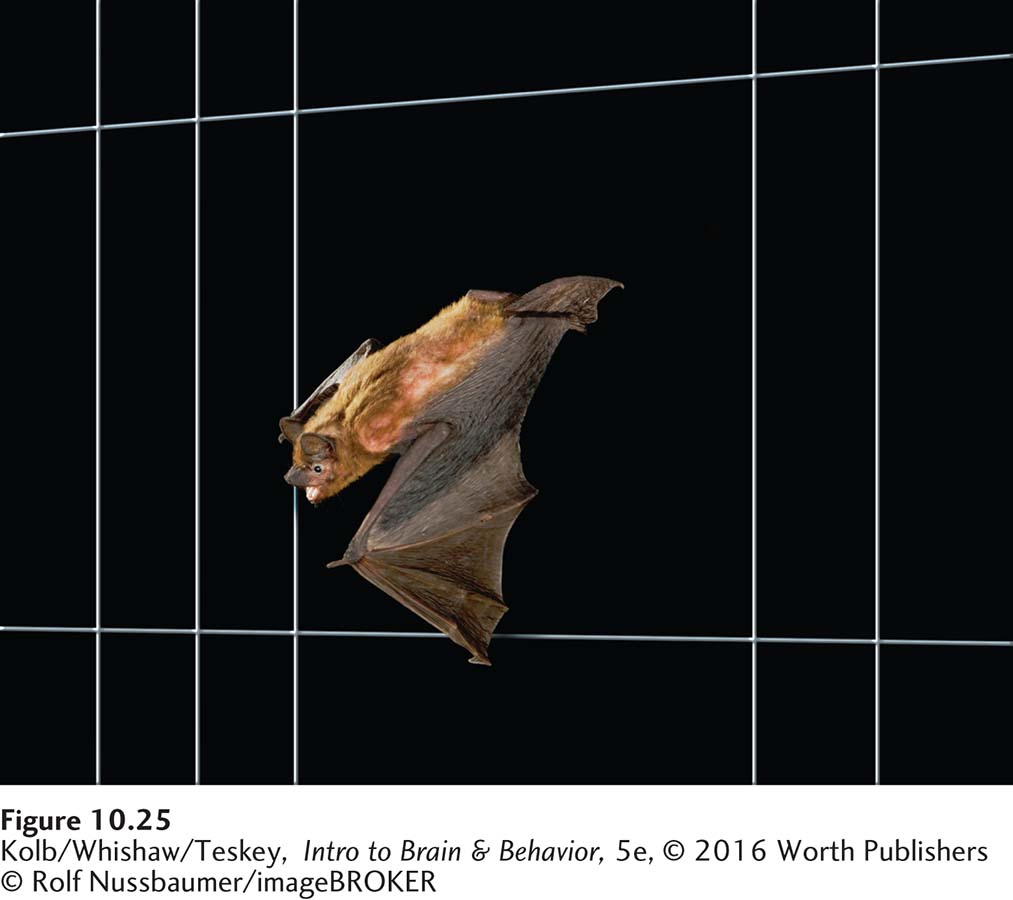
The echolocation abilities of bats are phenomenal, as shown in Figure 10-25. Bats in the wild can be trained to catch small food particles thrown up into the air in the dark. These echolocating skills make the bat a most efficient hunter. The little brown bat, for instance, can capture tiny flying insects, such as mosquitoes, at the remarkable rate of two per second.
Researchers have considerable interest in the neural mechanisms of bat echolocation. Each species emits sound waves in a relatively narrow frequency range, and a bat’s auditory pathway has cells specifically tuned to echoes in its species’ frequency range. For example, the mustached bat sends out sound waves ranging from 60,000 to 62,000 Hz, and its auditory system has a cochlear fovea (a maximally sensitive area in the organ of Corti) that corresponds to that frequency range.
Figure 9-2 details the fovea’s structure.
In this way, more neurons are dedicated to the frequency range used for echolocation than to any other range of frequencies. Analogously, our visual system dedicates more neurons to the retina’s fovea, the area responsible for our most detailed vision. In the cortex of the bat’s brain, several distinct areas process complex echoic inputs. One area computes the distance of given targets from the animal, for instance, whereas another area computes the velocity of a moving target. This neural system makes the bat exquisitely adapted for nighttime navigation.
10-5 REVIEW
Auditory Communication in Nonhuman Species
Before you continue, check your understanding.
Question 1
Song development in young birds is influenced both by genes and by early experience and learning, interactions indicative of ____________.
Question 2
In many bird species the control of song in the ____________ brain is lateralized to the hemisphere.
Question 3
Bats use ____________ to locate prey in the dark. This system is much like the ____________ that ships use to locate underwater objects.
Question 4
What does the presence of dialects in birdsong in the same species demonstrate?
Answers appear in the Self Test section of the book.