More than half of all described species are insects
During the Devonian period, more than 400 million years ago, some mandibulates colonized terrestrial environments. Of the several groups (including some crustacean isopods and decapods) that successfully colonized the land, none is more prominent today than the six-
As you saw at the opening of this chapter, more than 1 million of the 1.8 million described living species on Earth are insects, but that is a small fraction of the number of species of insects that are thought to exist on Earth. One of Terry Erwin’s experiments is shown in more detail in Investigating Life: How Many Unknown Species? This and similar experiments show that we have much to learn about the biodiversity of our own planet.
The wingless relatives of the insects—

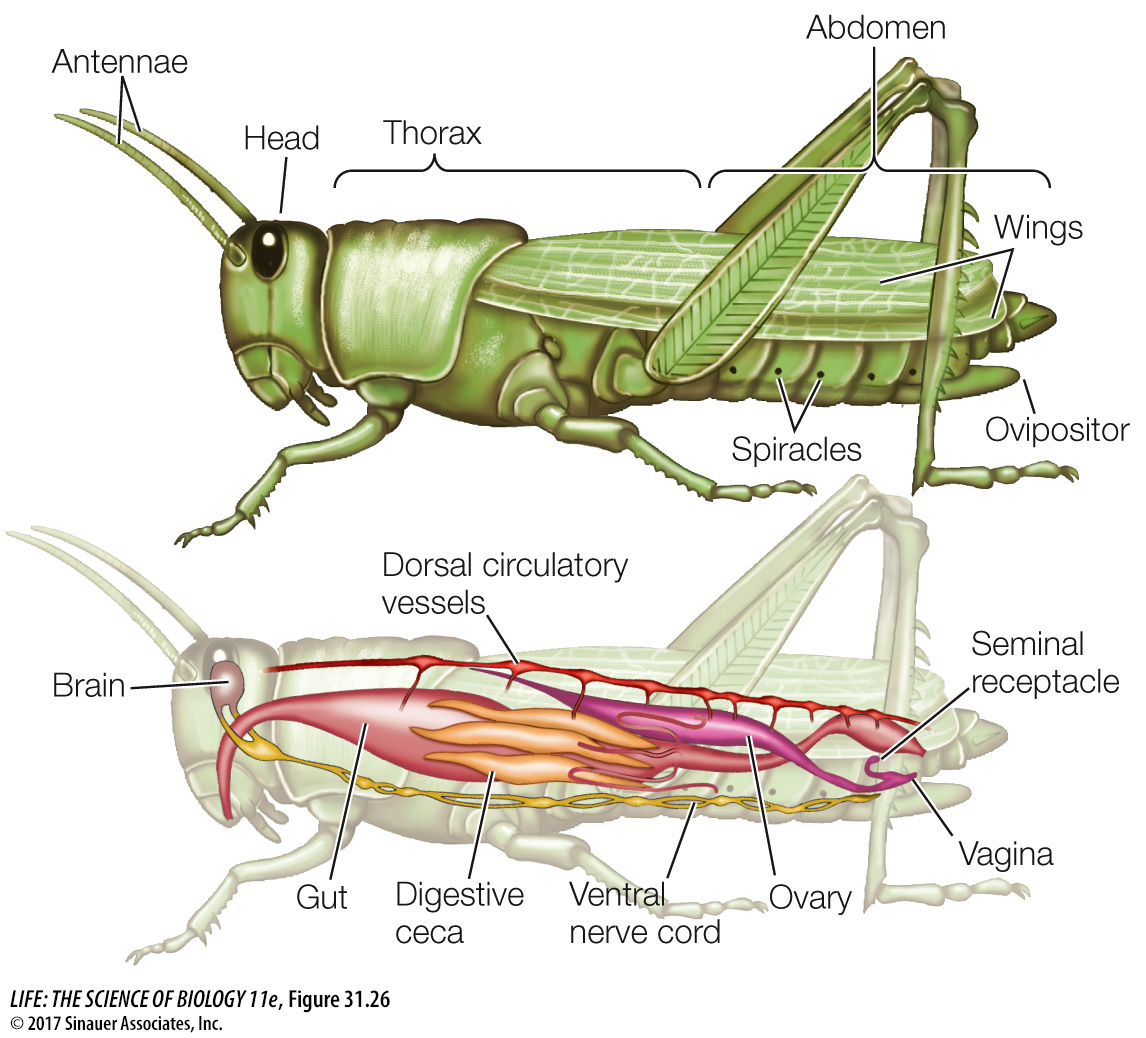
Like crustaceans, insects have a body with three regions: head, thorax, and abdomen. They have a single pair of antennae on the head and three pairs of legs attached to the thorax. In most groups of insects, the thorax also bears two pairs of wings. Unlike other arthropods, insects have no appendages growing from their abdominal segments (Figure 31.26). Insects are distinguished from springtails and other hexapods by their external mouthparts and by antennae that contain a motion-
investigating life
How Many Unknown Species?
experiment
Original Papers: Erwin, T. L. 1988. The tropical forest canopy: The heart of biotic diversity. In E. O. Wilson, ed., Biodiversity, pp. 123–
Erwin, T. L. 1997. Biodiversity at its utmost: Tropical forest beetles. In M. L. Reaka-
Entomologist Terry Erwin decided to estimate the number of undescribed species of insects. He devised and executed the following experiment in the forests of Panama.

work with the data
The data in the table were used by entomologist Terry Erwin to estimate the undescribed diversity of insects. Review the design of Erwin’s experiment above and in the opening story of this chapter. Then use Erwin’s data to answer the questions.
| Approximate number of beetle species collected from Luehea seemannii trees | 1,200 |
| Estimated number of host- |
163 |
| Number of tree species per hectare of Panamanian forest | 70 |
| Percent of beetle species living in tree canopy (as opposed to ground- |
75% |
| Percent of beetles among all insect species | 40% |
QUESTIONS
1.
From the data in the table, estimate the number of insect species in an average hectare of Panamanian forest. Assume that the data for beetles on L. seemannii are representative of the other tree species, and that all the species of beetles that are not host-
Estimates:
- Number of host-
specific beetles species in the forest canopy = (Number of beetles specific to L. seemannii) ´ (Number of species of canopy trees) = (163) × (70) = 11,410. - Number of non-
host- specific beetles species in the forest canopy = 1,200 – 163 = 1,037. - Number of beetle species on the forest floor = one-
third of the number of species in the canopy (75% of beetles are in the canopy, and 25% are on the forest floor). Based on (a) and (b), there are 11,410 + 1,037 = 12,447 species in a hectare of canopy. Therefore we can estimate 12,447/3 = 4,149 species of beetles on a hectare of forest floor. - Number of species of all insects other than beetles = 1.5 times the number of beetles (40% of insects are beetles, so 60% of insects are non-
beetles). From (a), (b), and (c), we can estimate 11,410 + 1,037 + 4,149 = 16,596 beetles in a hectare of Panamanian forest.
Therefore the number of insects other than beetles in a hectare of forest = (1.5) × (16,596) = 24,894.
This gives a total estimate for the number of insect species in an average hectare of Panamanian forest of 16,596 + 24,894 = 41,490.
2.
There are about 50,000 species of tropical forest trees. Assume that the data for beetles on L. seemannii are representative of other species of tropical trees and calculate the number of host-
- The estimate for the number of host-
specific tropical canopy insects = (50,000) × (163) = 8,150,000. - Add 1 million for generalist and temperate canopy beetles: total beetles = 8,150,000 + 1,000,000 = 9,500,000.
- As in Question 1, beetle species on the forest floor = one-
third of the number of species in the canopy. Therefore the number of species of ground beetles = 9,500,000/3 = 3,166,667 species. - As in Question 1, species of all insects other than beetles = 1.5 times the number of beetles. Our worldwide estimate for number of beetle species is 9,500,000 + 3,166,667 = 12,666,667. Therefore we estimate non-
beetle species diversity at (12,666,667) × (1.5) = 19,000,000. - Summing the number of species of beetles and non-
beetles, we get an estimate of worldwide insect species diversity of 12,666,667 + 19,000,000 = 31,666,667 insect species.
Note: Many biologists have debated whether Erwin’s assumptions are reasonable. Clearly, each estimate is highly dependent on how representative L. seemanii is as a tropical forest tree. If the average tropical forest tree has many fewer host-
Only about 1 million species of insects have been described by biologists worldwide to date (see Table 31.2). All entomologists agree that many more species of insects remain to be discovered, and many new species are discovered and described every year. Most entomologists currently think that Erwin’s estimates were high. Each of Erwin’s assumptions is now being tested; these tests require extensive work on additional species of trees, additional groups of insects, and in additional areas of the world.
A similar work with the data exercise may be assigned in LaunchPad.
Table 31.2 lists the major insect groups. Two groups—
| Group | Approximate number of described living species |
|---|---|
| Jumping bristletails (Archaeognatha) | 550 |
| Silverfish (Zygentoma) | 575 |
| PTERYGOTE (WINGED) INSECTS (PTERYGOTA) | |
| Mayflies (Ephemeroptera) | 3,250 |
| Dragonflies and damselflies (Odonata) | 6,000 |
| Neopterans (Neoptera)b | |
| Ice- |
35 |
| Gladiators (Mantophasmatodea) | 15 |
| Stoneflies (Plecoptera) | 3,800 |
| Webspinners (Embioptera) | 500 |
| Angel insects (Zoraptera) | 40 |
| Earwigs (Dermaptera) | 2,000 |
| Grasshoppers and crickets (Orthoptera) | 24,000 |
| Stick insects (Phasmida) | 3,000 |
| Cockroaches (Blattodea) | 4,500 |
| Termites (Isoptera) | 3,000 |
| Mantids (Mantodea) | 2,500 |
| Booklice and barklice (Psocoptera) | 5,750 |
| Thrips (Thysanoptera) | 6,000 |
| Lice (Phthiraptera) | 5,000 |
| True bugs, cicadas, aphids, leafhoppers (Hemiptera) | 104,000 |
| Holometabolous neopterans (Holometabola)c | |
| Ants, bees, wasps, sawflies (Hymenoptera) | 150,000 |
| Beetles (Coleoptera) | 390,000 |
| Strepsipterans (Strepsiptera) | 650 |
| Lacewings, ant lions, mantidflies (Neuroptera) | 6,000 |
| Dobsonflies, alderflies, fishflies (Megaloptera) | 350 |
| Snakeflies (Raphidoptera) | 250 |
| Scorpionflies (Mecoptera) | 800 |
| Fleas (Siphonaptera) | 2,000 |
| True flies (Diptera) | 125,000 |
| Caddisflies (Trichoptera) | 12,000 |
| Butterflies and moths (Lepidoptera) | 180,000 |
|
aThe hexapod relatives of insects include the springtails (Collembola; 3,000 spp.), two- bNeopteran insects can tuck their wings close to their bodies. cHolometabolous insects are neopterans that undergo complete metamorphosis. |
|
Hatchling pterygotes do not look like adults; they undergo substantial changes at each molt. The immature stages of insects between molts are called instars. A substantial change that occurs between one developmental stage and another is called metamorphosis. If the changes between stages are gradual, an insect is said to have incomplete metamorphosis. If the change between at least some stages is dramatic, an insect is said to have complete metamorphosis (see Figure 30.10). In many insects with complete metamorphosis, different stages are specialized for different functions and use different food sources. In many species the larvae are specialized for feeding and growing, whereas the adults are specialized for reproduction and dispersal.
Media Clip 31.8 Complete Metamorphosis
The adults of most flying insects have two pairs of stiff, membranous wings attached to the thorax. True flies, however, have one pair of wings and a pair of stabilizers called halteres (which are derived from the second pair of wings). In winged beetles, one pair of wings—
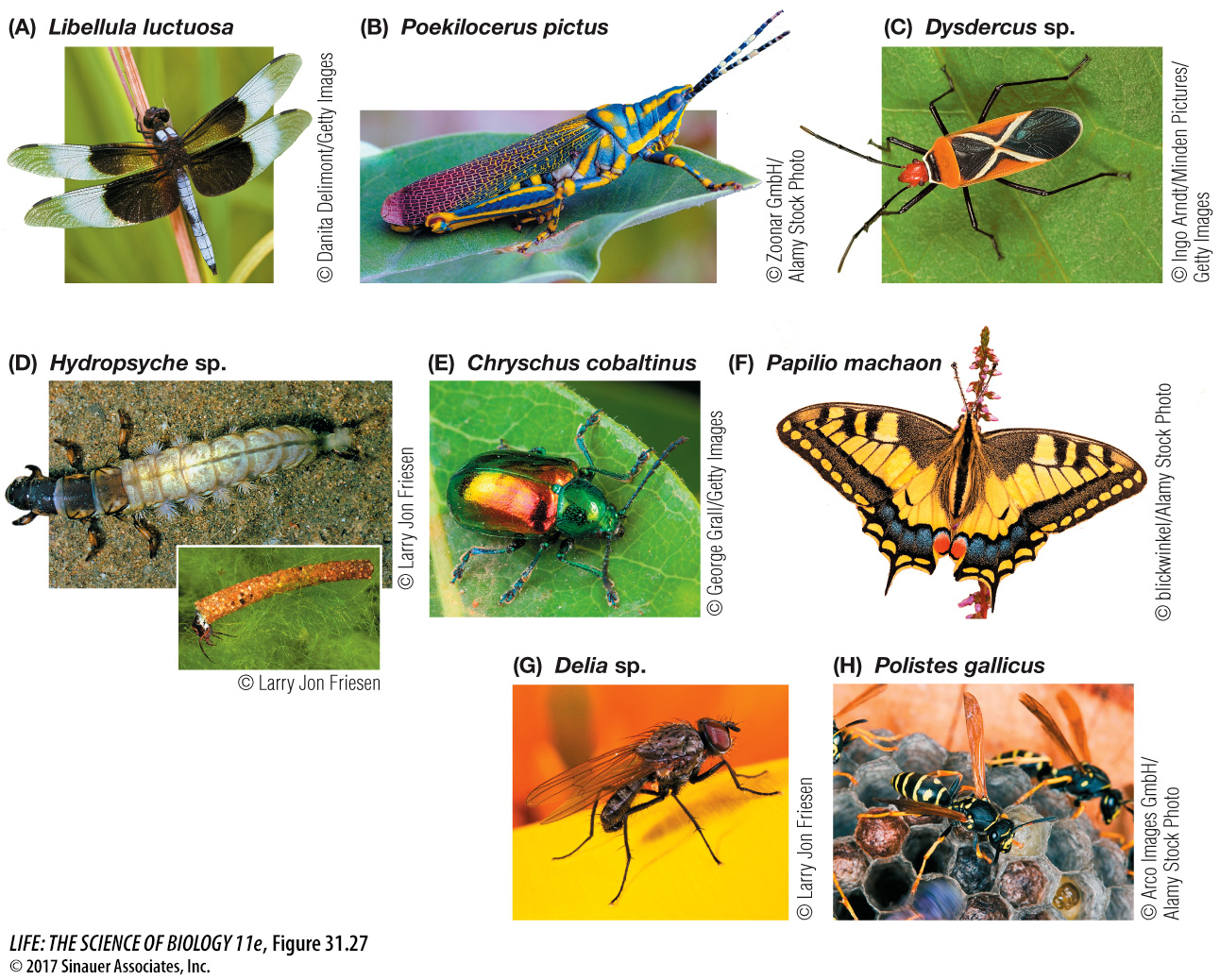
Two groups of pterygotes, the mayflies and dragonflies (Figure 31.27A), cannot fold their wings against their bodies. This is the ancestral condition for pterygote insects, and the mayflies and dragonflies are not closely related to one another. Members of both groups have predatory or herbivorous aquatic larvae that transform into flying adults after they crawl out of the water. Dragonflies (and their relatives the damselflies) are active predators as adults. In contrast, adult mayflies lack functional digestive tracts. Adult mayflies live only about a day, just long enough to mate and lay eggs.
All other pterygote insects—
More than 80 percent of all insects belong to a subgroup of the neopterans called the holometabolous insects (see Table 31.2), which undergo complete metamorphosis (Figure 31.27D). The many species of beetles account for almost half of this group (Figure 31.27E). Also included are lacewings and their relatives; caddisflies; butterflies and moths (Figure 31.27F); true flies (Figure 31.27G); and bees, wasps, and ants, some species of which display unique and highly specialized social behaviors (Figure 31.27H).
Molecular data suggest that insects began to diversify about 450 million years ago, about the time of the appearance of the first land plants. These early hexapods evolved in a terrestrial environment that lacked any other similar organisms, which in part accounts for their remarkable success. But the success of the insects is also due to their wings. Pterygote insects were the first animals in evolutionary history to achieve the ability to fly. Homologous genes control the development of insect wings and crustacean appendages, suggesting that the insect wing evolved from a dorsal branch of a crustacean-
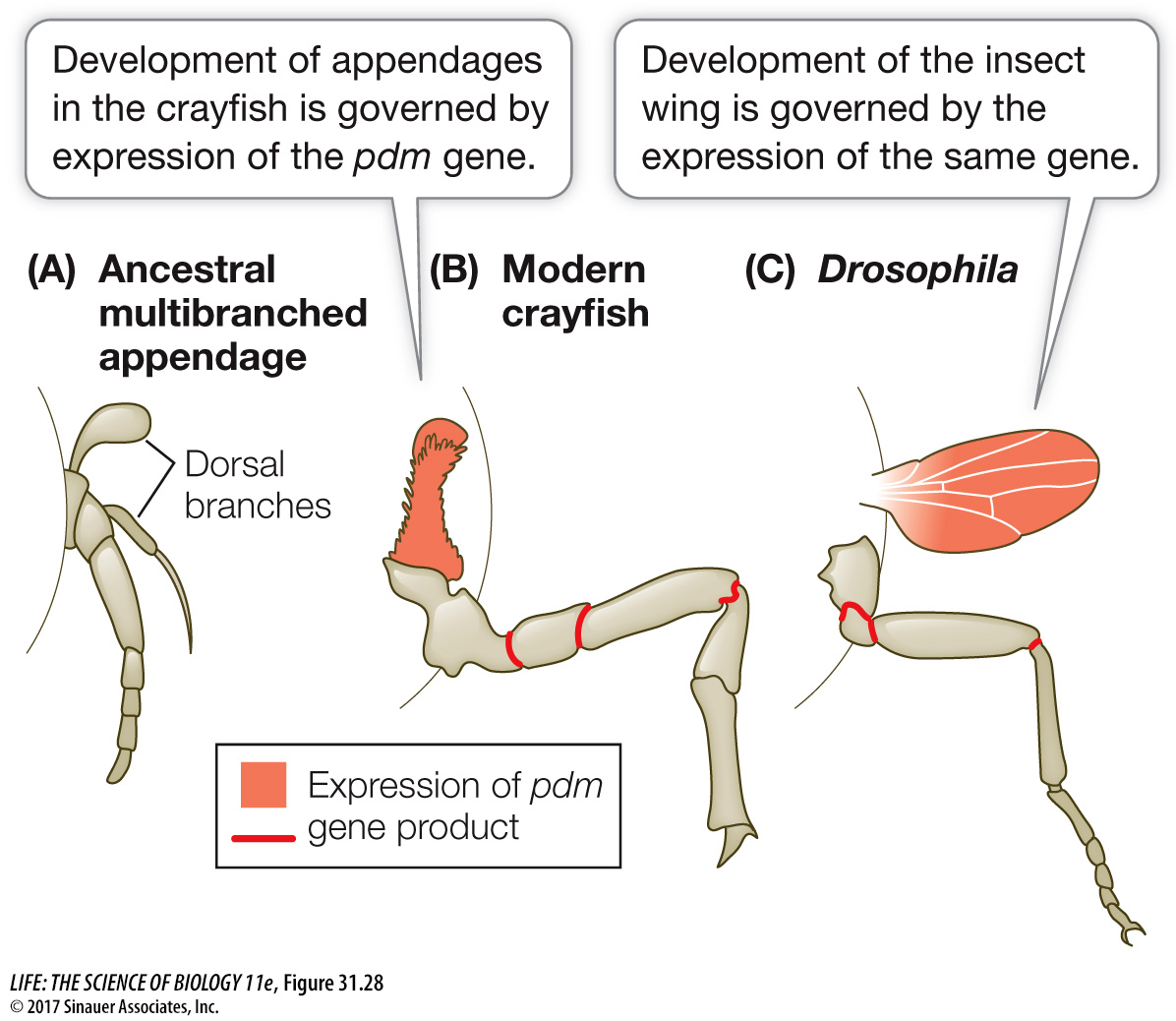
Q: If the hypothesized origin of insect wings is correct, then what was a possible function of the structure that would become a wing in flying insects?
The uppermost dorsal branch on the ancestral arthropod limb is thought to have functioned in gas exchange. The Hox gene expression data suggest that this structure may have been homologous to the insect wing.
Flight opened up many new lifestyles and feeding opportunities that only the insects could exploit, such as pollination of (and coevolution with) flowering plants. Flight is almost certainly one of the reasons for the remarkable numbers of both insect species and individual insects, and for their unparalleled evolutionary success.