Nuclear Exoribonucleases Degrade RNA That Is Processed Out of Pre-mRNAs
Because the human genome contains long introns, only about 5 percent of the nucleotides that are polymerized by RNA polymerase II during transcription are retained in mature, processed mRNAs. Although this process appears inefficient, it probably evolved in multicellular organisms because the process of exon shuffling facilitated the evolution of new genes in organisms with long introns (see Chapter 8). The introns that are spliced out and the RNA downstream from the cleavage/polyadenylation site are degraded by nuclear exoribonucleases.
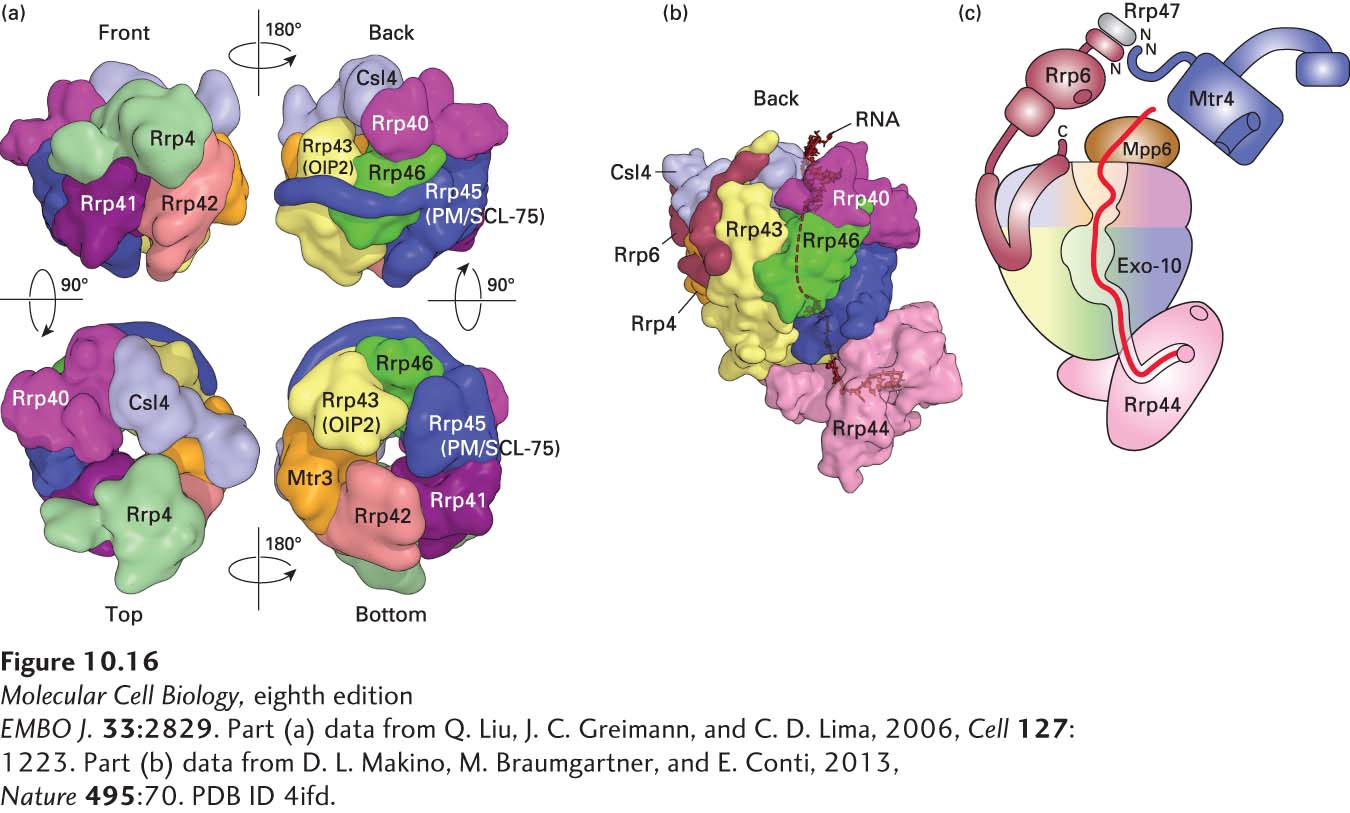
As mentioned earlier, the 2′,5′-phosphodiester bond in excised introns is hydrolyzed by a debranching enzyme (see Figure 10-11, step 6), yielding a linear molecule with unprotected ends. Such linear RNA molecules can be attacked by exoribonucleases, which hydrolyze one base at a time from the 5′ or 3′ end (as opposed to endoribonucleases, which digest internal phosphodiester bonds). The predominant mechanism of RNA decay is digestion by a large (~400-kDa) protein complex called the exosome, which contains an internal 3′→5′ exoribonuclease (Figure 10-16). (Exosomes also function in the cytoplasm, as discussed later.) The exosome is in many ways analogous to the proteasome (see Figure 3-31) that digests polyubiquitinylated proteins in both the nucleus and the cytoplasm. The predominant active site of the exosome lies on the inside of the complex, where it can digest only single-stranded RNAs that are threaded into the pore at the top of the complex (Figure 10-16b). This pore is too small to allow the entry of double-stranded or other structured regions of RNAs. Other proteins that associate with the complex include an RNA helicase, which disrupts base pairing and RNA-protein interactions that would otherwise prevent the entry of RNA into the pore.
EMBO J.33:2829. [Part (a) data from Q. Liu, J. C. Greimann, and C. D. Lima, 2006, Cell127:1223. Part (b) data from D. L. Makino, M. Braumgartner, and E. Conti, 2013, Nature495:70. PDB ID 4ifd.]
FIGURE 10-16Structure of the exosome. (a) Catalytically inactive exosome core. A nine-subunit, 286-kDa human exosome core was assembled in vitro from subunits Rrp41, Rrp45, Rrp42, Mtr3, Rrp43, Rrp46, Rrp4, Rrp40, and Csl4 expressed at a high level in E. coli (see Figure 6-29). Its structure was determined to a resolution of 3.35 Å by x-ray crystallography. (b) The 10-subunit, catalytically active cytoplasmic exosome. The orientation is similar to that of the upper right image in part (a), but rotated slightly counterclockwise. Processive 3′→5′ exonuclease activity is provided by the tenth subunit, Rrp44 (pink), associated with the bottom of the core. The C-terminus of an eleventh subunit, Rrp6, in the nuclear exosome is shown in maroon. RNA with a double-stranded region at the top and a 3′ single-stranded region that enters the core pore is shown in black. (c) Diagram of the 14-subunit nuclear exosome. Exo-10 represents the 10-subunit complex shown in (b). A heterodimer of Rrp6 and Rrp47 associates with Csl4 at the top of the exosome core through the C-terminal domain of Rrp6, as shown in (b). The N-terminus of an RNA helicase, Mtr4 (blue), associates with the heterodimerization domain of Rrp6 and Rrp47. Another subunit associated with the top, Mpp6, also associates with the Mtr4 RNA helicase in the human nuclear exosome, but its structure and the details of the Mpp6-Mtr4 interaction remain to be determined. The path of single-stranded RNA through the exosome is diagrammed in red. The exonuclease active site in the processive exonuclease Rrp44 is indicated by a pink circle. An endonuclease active site in Rrp44 is represented by the pink oval. A non-processive 3′→5′ exonuclease active site in Rrp6 is represented by a maroon oval. See B. Schuch et al., 2014.
EMBO J.33:2829. [Part (a) data from Q. Liu, J. C. Greimann, and C. D. Lima, 2006, Cell127:1223. Part (b) data from D. L. Makino, M. Braumgartner, and E. Conti, 2013, Nature495:70. PDB ID 4ifd.]
In addition to introns, the exosome also degrades pre-mRNAs that have not been properly spliced or polyadenylated, although at present, it is not yet clear how the exosome recognizes improperly processed pre-mRNAs. But in yeast cells with temperature-sensitive mutant PAP (see Figure 10-15), pre-mRNAs are retained at their sites of transcription in the nucleus at the nonpermissive temperature. These abnormally processed pre-mRNAs are released in cells with a second mutation in a subunit of the exosome found only in nuclear and not in cytoplasmic exosomes (Rrp6, see Figure 10-16). In addition, exosomes are found concentrated at sites of transcription in Drosophila polytene chromosomes, where they are associated with RNA polymerase II elongation factors. These results suggest that the exosome participates in an as yet poorly understood quality-control mechanism in the nucleus that recognizes aberrantly processed pre-mRNAs, preventing their export to the cytoplasm and ultimately leading to their degradation.
To avoid being degraded by nuclear exonucleases, nascent transcripts, pre-mRNA-processing intermediates, and mature mRNAs in the nucleus must have their ends protected. As discussed above, the 5′ end of a nascent transcript is protected by addition of the 5′ cap structure as soon as the 5′ end emerges from the polymerase. The 5′ cap is protected because it is bound by a heterodimeric nuclear cap-binding complex (CBC), which protects it from 5′ exonucleases and also functions in export of the mRNA to the cytoplasm. The 3′ end of a nascent transcript lies within the RNA polymerase and is thus inaccessible to exonucleases (see Figure 5-12). As discussed previously, the free 3′ end generated by cleavage of a pre-mRNA downstream from the polyadenylation signal is rapidly polyadenylated by the PAP associated with the other 3′ processing factors, and the resulting poly(A) tail is bound by PABPN1 (see Figure 10-15). This tight coupling of cleavage and polyadenylation, followed by PABPN1 binding, protects the 3′ end from exonuclease attack.