Splicing Repressors and Activators Control Splicing at Alternative Sites
As is evident from Figure 10-18, the Drosophila Sxl protein and Tra protein have opposite effects: Sxl prevents splicing, causing exons to be skipped, whereas Tra promotes splicing. The action of similar proteins may explain the cell-type-specific expression of fibronectin isoforms in humans (see Figure 5-16). For instance, an Sxl-like splicing repressor expressed in hepatocytes might bind to splice sites for the EIIIA and EIIIB exons in the fibronectin pre-mRNA, causing them to be skipped during RNA splicing. Alternatively, a Tra-like splicing activator expressed in fibroblasts might activate the splice sites associated with those exons, leading to their inclusion in the mature mRNA. Experimental examination of some systems has revealed that the inclusion of an exon in some cell types and the skipping of the same exon in other cell types results from the combined influence of several splicing repressors (usually hnRNP proteins) and enhancers (usually SR proteins). RNA binding sites for repressors can also occur in exons, where they are called exonic splicing silencers. And binding sites for splicing activators can also occur in introns, where they are called intronic splicing enhancers.
Alternative splicing of exons is especially common in the nervous system, where it generates multiple isoforms of many proteins required for neuronal development and function in both vertebrates and invertebrates. The primary transcripts of the genes encoding these proteins often show complex splicing patterns that can generate several different mRNAs, which are expressed in different anatomic locations within the central nervous system. Here we consider two remarkable examples that illustrate the critical role of this process in neural function.
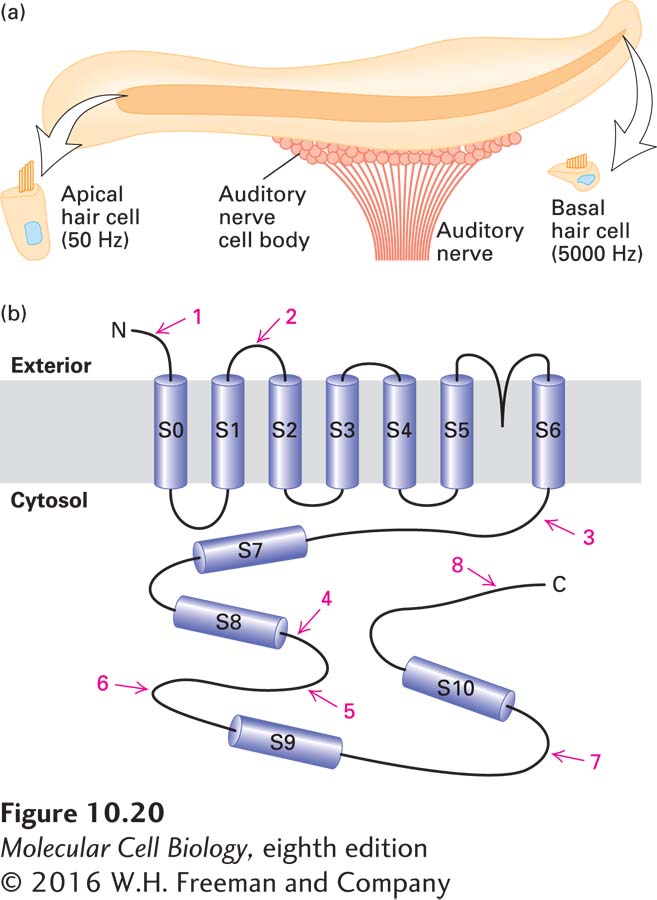
Expression of K+-Channel Proteins in Vertebrate Hair Cells In the inner ear of vertebrates, individual hair cells, which are ciliated neurons, respond most strongly to a specific frequency of sound. Cells tuned to low frequencies (~50 Hz) are found at one end of the tubular cochlea that makes up the inner ear; cells responding to high frequencies (~5000 Hz) are found at the other end (Figure 10-20a). Cells in between the two ends respond to a gradient of frequencies between these extremes. One component in the tuning of hair cells in reptiles and birds is the opening of K+ ion channels in response to increased intracellular Ca2+ concentrations. The Ca2+ concentration at which the channel opens determines the frequency with which the membrane potential oscillates and hence the frequency to which the cell is tuned.
FIGURE 10-20Role of alternative splicing in the perception of sounds of different frequencies. (a) The chicken cochlea, a 5-mm-long tube, contains an epithelium of auditory hair cells that are tuned to a gradient of vibrational frequencies from 50 Hz at the apical end (left) to 5000 Hz at the basal end (right). (b) The Ca2+-activated K+ channel contains seven transmembrane α helices (S0–S6), which associate to form the channel. The cytosolic domain, which includes four hydrophobic regions (S7–S10), regulates opening of the channel in response to Ca2+. Isoforms of the channel, encoded by alternatively spliced mRNAs produced from the same primary transcript, open at different Ca2+ concentrations and thus respond to different frequencies. Red numbers refer to regions where alternative splicing produces different amino acid sequences in the various isoforms. See K. P. Rosenblatt et al., 1997, Neuron19:1061.
The gene encoding this Ca2+-activated K+ channel is expressed as multiple, alternatively spliced mRNAs, which encode proteins that open at different Ca2+ concentrations. Hair cells with different response frequencies express different isoforms of the channel protein depending on their position along the length of the cochlea. The sequence variation in the protein is very complex: there are at least eight regions in the mRNA where one of several alternative exons is utilized, permitting the expression of 576 possible isoforms (Figure 10-20b). PCR analysis of mRNAs from individual hair cells has shown that each hair cell expresses a mixture of different K+-channel mRNAs, with different isoforms predominating in different cells according to their position along the cochlea. This remarkable arrangement suggests that splicing of the K+-channel pre-mRNA is regulated in response to extracellular signals that inform the cell of its position along the cochlea.
Other studies have demonstrated that splicing at one of the alternative splice sites in the Ca2+-activated K+-channel pre-mRNA in the rat is suppressed when a specific protein kinase is activated by neuron depolarization in response to synaptic activity from interacting neurons. This observation raises the possibility that a splicing repressor specific for this splice site may be activated when it is phosphorylated by this protein kinase, whose activity in turn is regulated by synaptic activity. Since hnRNP and SR proteins are extensively modified by phosphorylation and other post-translational modifications, it seems likely that complex regulation of alternative RNA splicing through post-translational modifications of splicing factors plays a significant role in modulating neuron function.
Page 438
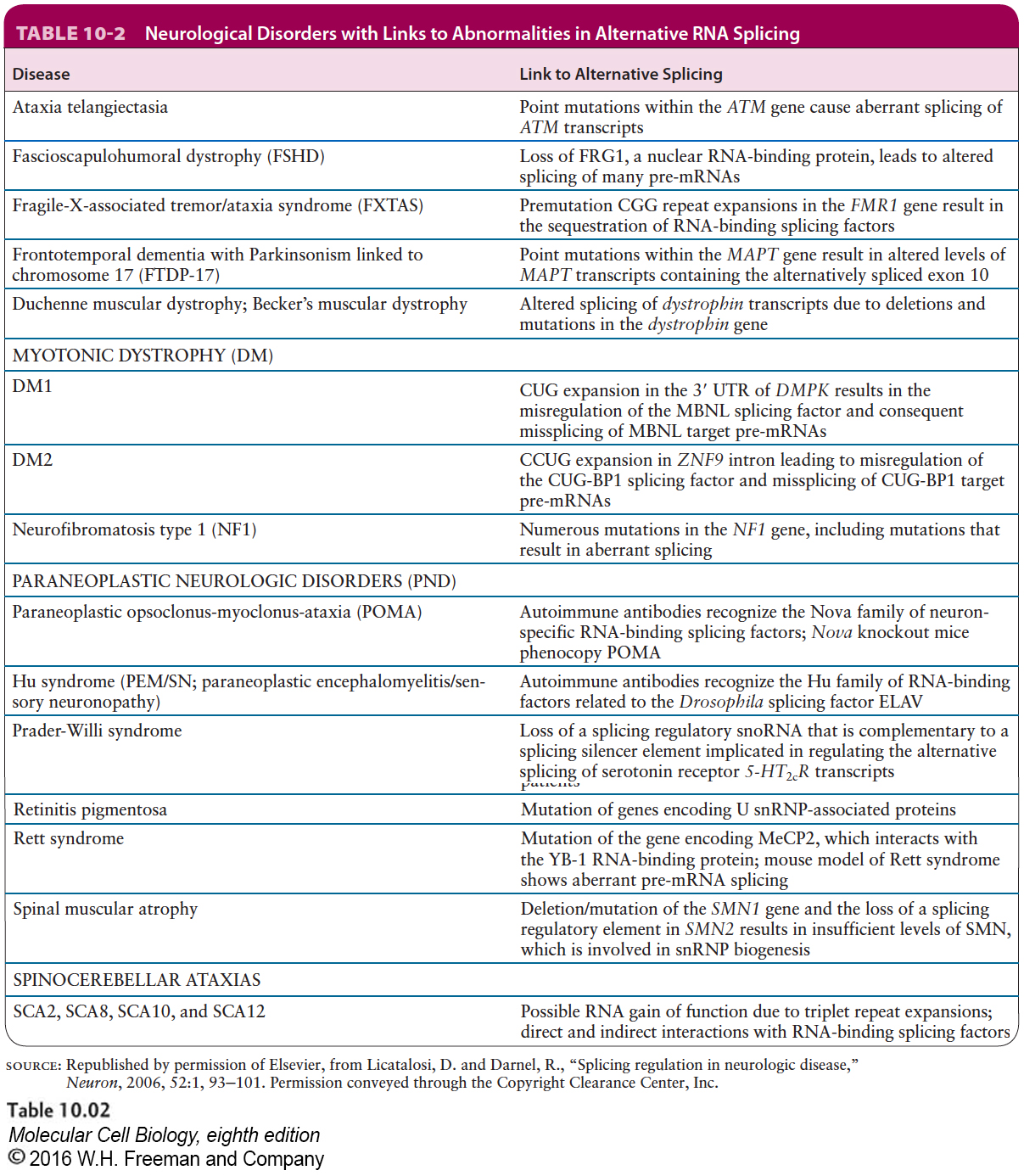
Many examples of genes similar to those that encode the cochlear K+ channel have been observed in vertebrate neurons; in these cases, alternatively spliced mRNAs co-expressed from a specific gene in one type of neuron are expressed at different relative concentrations in different regions of the central nervous system. Expansions in the number of microsatellite repeats within the transcribed regions of genes expressed in neurons can alter the relative concentrations of alternatively spliced mRNAs transcribed from multiple genes. In Chapter 8, we discussed how backward slippage during DNA replication can lead to expansion of a microsatellite repeat (see Figure 8-5). At least 14 different types of neurological diseases result from expansion of microsatellite regions within transcription units expressed in neurons. The resulting long regions of repeated simple sequences in nuclear pre-mRNAs of these neurons result in abnormalities in the relative concentrations of alternatively spliced mRNAs. For example, the most common of these types of diseases, myotonic dystrophy, results from increased copies of either CUG repeats in one transcript, in some patients, or CCUG repeats in another transcript, in other patients. When the number of these repeats increases to 10 or more times the normal number of repeats, abnormalities are observed in the functions of two hnRNP proteins that bind to these repeated sequences. The abnormalities probably result because the hnRNPs are bound by the abnormally high concentrations of the repeats in the nuclei of neurons in these patients and cannot associate with other pre-mRNAs. This sequestration of the hnRNPs leads to alterations in the rate of splicing of different alternative splice sites in multiple pre-mRNAs that are normally regulated by these hnRNP proteins. Because of the importance of the proper regulation of alternative splicing for the normal function of neurons, multiple human neurological disorders are associated with abnormalities in the function of nuclear RNA-binding proteins and the expansion of microsatellite repeats that generate binding sites for splicing factors (Table 10-2).
Page 439
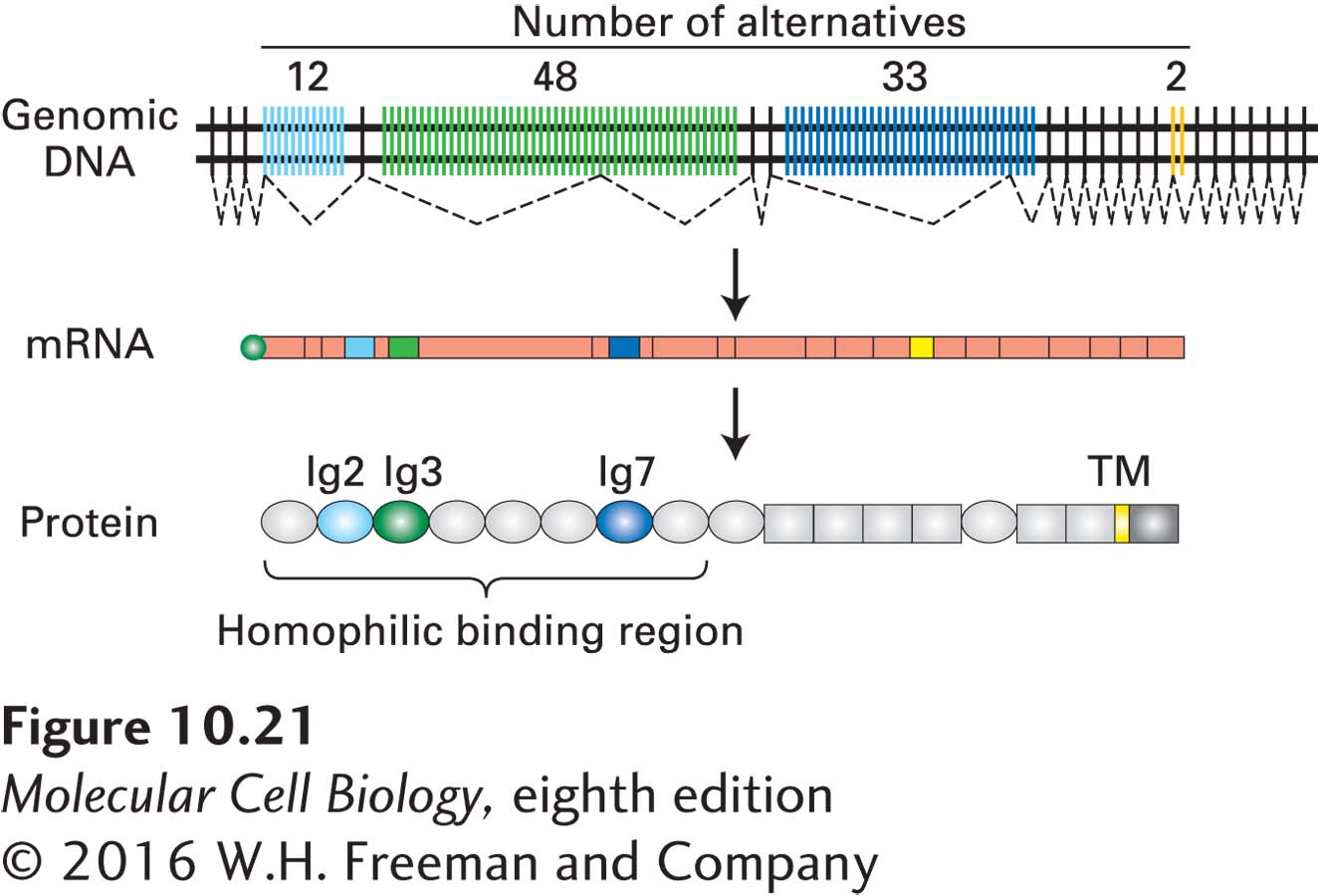
Expression of Dscam Isoforms in Drosophila Retinal Neurons The most extreme example of regulated alternative RNA processing yet uncovered occurs in expression of the Dscam gene in Drosophila. Mutations in this gene interfere with the normal synaptic connections made between retinal axons and dendrites during fly development. Analysis of the Dscam gene showed that it contains four groups of exons within which one of several possible exons is included in the final mature mRNA. The gene contains a total of 95 exons (Figure 10-21), generating 38,016 possible alternatively spliced isoforms! Drosophila mutants with a version of the gene that can be spliced in only about 22,000 different ways have specific defects in connectivity between neurons. These results indicate that expression of most of the possible Dscam isoforms through regulated RNA splicing helps to specify the tens of millions of different specific synaptic connections between neurons in the Drosophila brain. In other words, the correct wiring of neurons in the brain requires regulated RNA splicing.
FIGURE 10-21The Drosophila Dscam gene is processed into a vast number of alternative isoforms.Dscam encodes a cell-surface protein on neurons. The protein (bottom) is composed of ten different immunoglobulin (Ig) domains (ovals), six different fibronectin type III domains (rectangles), one transmembrane domain (yellow), and a C-terminal cytoplasmic domain (dark gray). The fully processed mRNA is shown as rectangles representing each exon, with the length of the rectangle corresponding to the length of the exons, and a green circle representing the 5’ cap. Each mRNA contains one of the 12 Ig2 exons shown in light blue (top), one of the 48 Ig3 exons shown in green, one of the 33 Ig7 exons shown in dark blue, and one of the 2 transmembrane exons shown in yellow. The exons shown in pink are spliced into each of the messages. Thus alternative splicing can generate 12 × 48 × 33 × 2 = 38,016 possible isoforms. See M. R. Sawaya et al., 2008, Cell134:1007.