HIV Rev Protein Regulates the Transport of Unspliced Viral mRNAs
As discussed earlier, transport of mRNPs containing mature, functional mRNAs through NPCs from the nucleus to the cytoplasm entails a complex mechanism that is crucial to gene expression (see Figures 10-23, 10-24, and 10-25). Regulation of this transport theoretically could provide another means of gene control, although it appears to be relatively rare. Indeed, the only known examples of regulated mRNA export occur during the cellular response to conditions (e.g., heat shock) that cause protein denaturation or during viral infection, when virus-induced alterations in nuclear export of mRNPs maximize viral replication. Here we describe the regulation of mRNP export mediated by a protein encoded by human immunodeficiency virus (HIV).
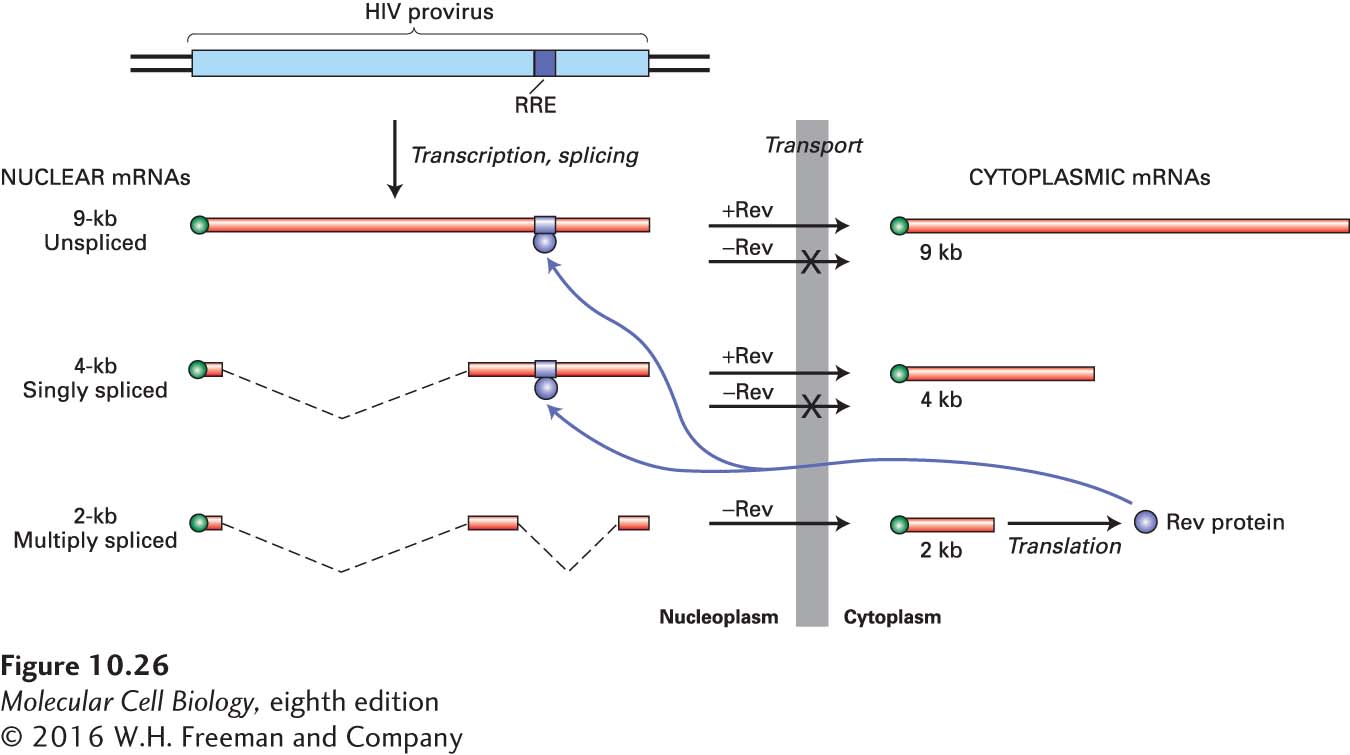
HIV, which is a retrovirus, integrates a DNA copy of its RNA genome into the host-cell DNA (see Figure 5-48). The integrated viral DNA, or provirus, contains a single transcription unit, which is transcribed into a single primary transcript by cellular RNA polymerase II. The HIV transcript can be spliced in alternative ways to yield three classes of mRNAs: a 9-kb unspliced mRNA; 4-kb mRNAs formed by removal of one intron; and 2-kb mRNAs formed by removal of two or more introns (Figure 10-26). After their synthesis in the host-cell nucleus, all three classes of HIV mRNAs are transported to the cytoplasm and translated into viral proteins; some of the 9-kb unspliced RNA is used as the viral genome in progeny virions that bud from the cell surface.
FIGURE 10-26Transport of HIV mRNAs from the nucleus to the cytoplasm. The HIV genome, which contains several coding regions, is transcribed into a single 9-kb primary transcript. Several 4-kb mRNAs result from the splicing out of any one of several introns (dashed lines), and several 2-kb mRNAs result from the splicing out of two or more alternative introns. After transport to the cytoplasm, these various RNA species are translated into different viral proteins. Rev protein, encoded by a 2-kb mRNA, interacts with the Rev-response element (RRE) in the unspliced and singly spliced mRNAs, stimulating their transport to the cytoplasm. See B. R. Cullen and M. H. Malim, 1991, Trends Biochem. Sci.16:346.
Since the 9-kb and 4-kb HIV mRNAs contain splice sites, they can be viewed as incompletely spliced mRNAs. As discussed earlier, association of such incompletely spliced mRNAs with snRNPs in spliceosomes normally blocks their export from the nucleus. Thus HIV, as well as other retroviruses, must have some mechanism for overcoming this block, permitting export of the longer viral mRNAs. Some retroviruses have evolved an RNA sequence within their genome called the constitutive transport element (CTE), which binds to the NXF1/NXT1 mRNP exporter with high affinity. This strong interaction with the mRNP exporter allows export of unspliced retroviral RNA into the cytoplasm. HIV solved the problem differently.
Studies with HIV mutants showed that transport of unspliced 9-kb and singly spliced 4-kb viral mRNAs from the nucleus to the cytoplasm requires the virus-encoded Rev protein. Subsequent biochemical experiments demonstrated that Rev binds to a specific Rev-response element (RRE) that is present in HIV RNA. In cells infected with HIV mutants lacking the RRE, unspliced and singly spliced viral mRNAs remain in the nucleus, demonstrating that the RRE is required for Rev-mediated stimulation of nuclear export. Early in an infection, before any Rev protein is synthesized, only multiply spliced 2-kb mRNAs that do not retain any splice sites can be exported. One of these alternatively spliced 2-kb mRNAs encodes Rev, which contains a leucine-rich nuclear-export signal that interacts with the transporter exportin 1 (see Chapter 13) rather than with the NXF1/NXT1 mRNP exporter. Translation of Rev in the cytoplasm, followed by its import into the nucleus, results in export of the larger unspliced and singly spliced HIV mRNAs through the NPC.