CHAPTER13:Moving Proteins into Membranes and Organelles
[From Wei, D. et al., “High-resolution three-dimensional reconstruction of a whole yeast cell using focused-ion beam scanning electron microscopy,” Biotechniques, 2012, 53(1):41–48.]
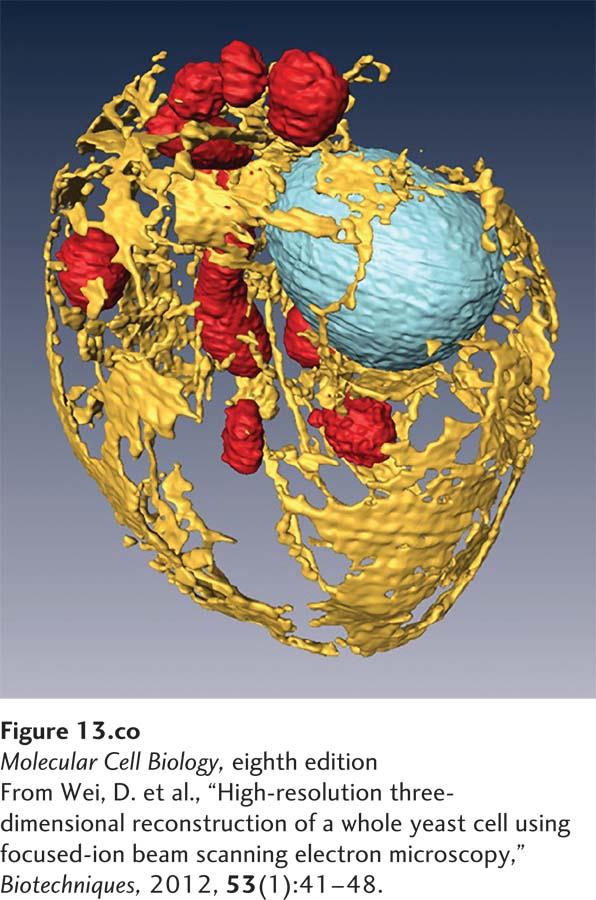
A three-dimensional reconstruction of the internal membranes of a yeast cell using scanning electron microscopy. The cell wall has been removed and the organelles highlighted with false color to reveal the endoplasmic reticulum (yellow), mitochondria (red), and nucleus (blue). Cell diameter is 3.5 µm.
[From Wei, D. et al., “High-resolution three-dimensional reconstruction of a whole yeast cell using focused-ion beam scanning electron microscopy,” Biotechniques, 2012, 53(1):41–48.]
A typical mammalian cell contains up to 10,000 different kinds of proteins; a yeast cell, about 5000. The vast majority of these proteins are synthesized by cytosolic ribosomes, and many remain within the cytosol (see Chapter 5). However, as many as half of the different kinds of proteins produced in a typical cell are delivered to one or another of the various membrane-bounded organelles within the cell or to the cell surface. For example, many receptor proteins and transport proteins must be delivered to the plasma membrane, some water-soluble enzymes such as RNA and DNA polymerases must be targeted to the nucleus, and components of the extracellular matrix, as well as digestive enzymes and polypeptide signaling molecules, must be directed to the cell surface for secretion from the cell. These and all the other proteins produced by a cell must reach their correct locations for the cell to function properly.
The delivery of newly synthesized proteins to their proper cellular destinations, usually referred to as protein targeting or protein sorting, encompasses two very different kinds of processes: signal-based targeting and vesicle-based trafficking. The first kind of process involves the targeting of a newly synthesized protein from the cytoplasm to an intracellular organelle. Targeting can occur during translation or soon after synthesis of the protein is complete. For membrane proteins, targeting leads to insertion of the protein into the lipid bilayer of the membrane, whereas for water-soluble proteins, targeting leads to translocation of the entire protein across the membrane into the aqueous interior of the organelle. Proteins are sorted to the endoplasmic reticulum (ER), mitochondria, chloroplasts, peroxisomes, and nucleus by this general process (Figure 13-1).
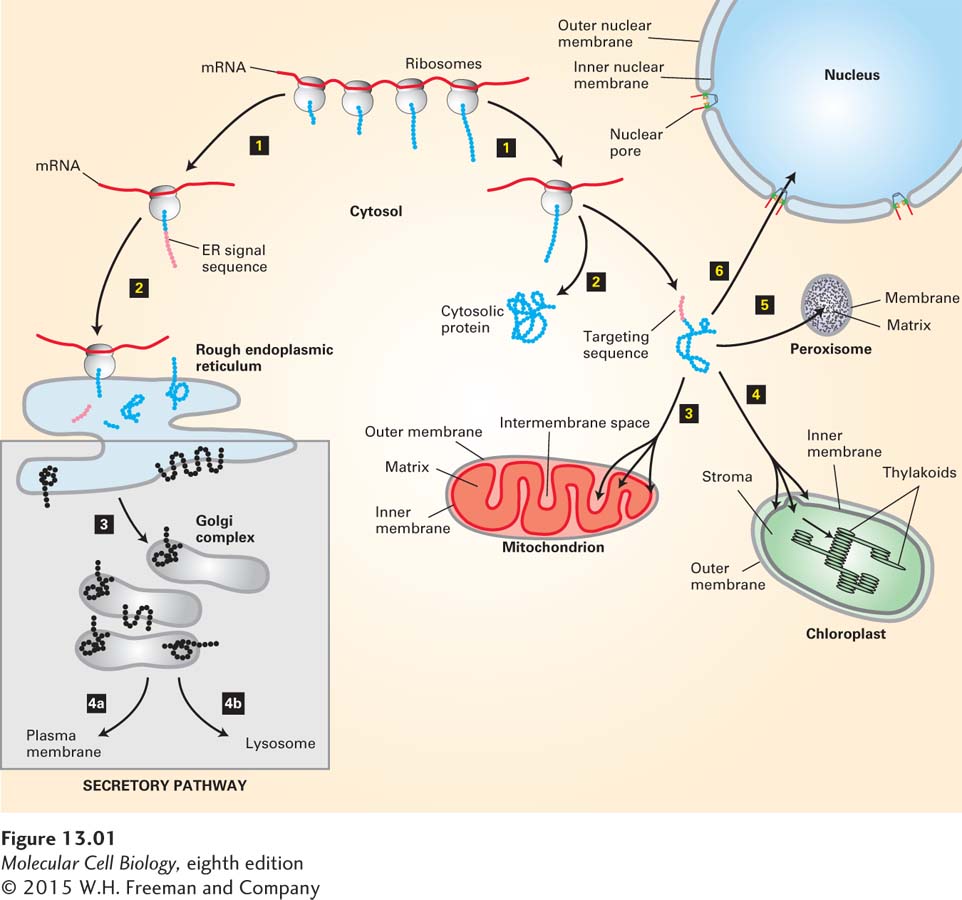
FIGURE 13-1Overview of major protein-sorting pathways in eukaryotes. All nuclear DNA–encoded mRNAs are translated on cytosolic ribosomes. Right (nonsecretory pathways): Synthesis of proteins lacking an ER signal sequence is completed on free ribosomes (step 1). Those proteins that contain no targeting sequence are released into the cytosol and remain there (step 2). Proteins with an organelle-specific targeting sequence (pink) are first released into the cytosol (step 2) but are then imported into mitochondria, chloroplasts, peroxisomes, or the nucleus (steps 3–6). Mitochondrial and chloroplast proteins typically pass through the outer and inner membranes to enter the matrix or stromal space, respectively. Other proteins are sorted to other subcompartments of these organelles by additional sorting steps. Nuclear proteins enter and exit through visible pores in the nuclear envelope. Left (secretory pathway): Ribosomes synthesizing nascent proteins in the secretory pathway are directed to the rough endoplasmic reticulum (ER) by an ER signal sequence (pink; steps 1 and 2). After translation is completed on the ER, these proteins can move via transport vesicles to the Golgi complex (step 3). Further sorting delivers proteins either to the plasma membrane or to lysosomes (step 4a or 4b). The vesicle-based processes underlying the secretory pathway (steps 3 and 4, shaded box) are discussed in Chapter 14.
Page 584
The second general sorting process, known as the secretory pathway, involves transport of proteins from the ER to their final destination within membrane-enclosed vesicles. For many proteins, including those that make up the extracellular matrix, the final destination is the outside of the cell (hence the name); integral membrane proteins are also transported to the Golgi complex, lysosomes, and plasma membrane by this process. The secretory pathway begins in the ER; thus all proteins slated to enter the secretory pathway are initially targeted to this organelle.
Targeting to the ER usually involves nascent proteins still in the process of being synthesized on a ribosome. Newly made proteins are thus extruded from the ribosome directly into the ER membrane. Once translocated across the ER membrane, proteins are assembled into their native conformation by protein-folding catalysts present in the lumen of the ER. Indeed, the ER is the location where about one-third of the proteins in a typical cell fold into their native conformations, and most of the resident ER proteins either directly or indirectly contribute to the folding process. As part of the folding process, proteins also undergo specific post-translational modifications in the ER. These processes are monitored carefully, and only after their folding and assembly is complete are proteins permitted to be transported out of the ER to other destinations. Proteins whose final destination is the Golgi complex, lysosomes, plasma membrane, or cell exterior are transported along the secretory pathway by the action of small vesicles that bud from the membrane of one organelle and then fuse with the membrane of another (see Figure 13-1, shaded box). We discuss vesicle-based protein trafficking in the next chapter because mechanistically it differs significantly from non-vesicle-based protein targeting to intracellular organelles.
Page 585
In this chapter, we examine how proteins are targeted to five intracellular organelles: the ER, mitochondria, chloroplasts, peroxisomes, and nucleus. Two features of this protein-targeting process were initially quite baffling: how a given protein could be directed to only one specific membrane, and how relatively large hydrophilic protein molecules could be translocated across a hydrophobic membrane without disrupting the function of the bilayer as a barrier to ions and small molecules. Using a combination of biochemical purification methods and genetic screens for identifying mutants unable to execute particular translocation steps, cell biologists have identified many of the cellular components required for translocation across each of the different intracellular membranes. In addition, many of the major translocation processes in the cell have been reconstituted by incorporating their purified protein components into artificial lipid bilayers. Such in vitro systems can be freely manipulated experimentally.
These studies have shown that, despite some variations, the same basic mechanisms govern protein sorting to all the various intracellular organelles. We now know, for instance, that the information to target a protein to a particular organelle destination is encoded within the amino acid sequence of the protein itself, usually within a sequence of about 20 amino acids, known generically as a targeting sequence (see Figure 13-1); these sequences are also called signal sequences or signal peptides. Such targeting sequences are usually located at the N-terminus of a protein and are thus the first part of the protein to be synthesized. More rarely, targeting sequences are located at either the C-terminus or within the interior of a protein sequence. Each organelle carries a set of receptor proteins that bind (directly or indirectly) only to specific kinds of targeting sequences, thus ensuring that the information encoded in a targeting sequence governs the specificity of targeting. Once a protein containing a targeting sequence has interacted with the corresponding receptor, the polypeptide chain is transferred to some kind of translocation channel that allows the protein to pass into or through the membrane bilayer. The unidirectional transfer of a protein into an organelle, without its sliding back out into the cytoplasm, is usually achieved by coupling translocation to an energetically favorable process such as hydrolysis of GTP or ATP. Some proteins are subsequently sorted further to reach a subcompartment within the target organelle; such sorting depends on yet other signal sequences and other receptor proteins. Finally, targeting sequences may be removed from the mature protein by specific proteases.
For each of the protein-targeting events discussed in this chapter, we will seek to answer four fundamental questions:
What is the nature of the targeting sequence, and what distinguishes it from other types of targeting sequences?
What is the receptor for the targeting sequence?
What is the structure of the translocation channel that allows transfer of proteins across the membrane bilayer? In particular, is the channel so narrow that proteins can pass through only in an unfolded state, or will it accommodate folded protein domains?
What is the source of energy that drives unidirectional transfer across the membrane?
In the first part of the chapter, we cover targeting of proteins to the ER, including the post-translational modifications that proteins undergo as they enter the secretory pathway. Targeting of proteins to the ER is the best-understood example of protein targeting and will serve as an exemplar of the process in general. We then describe targeting of proteins to mitochondria, chloroplasts, and peroxisomes. Finally, we cover the transport of proteins into and out of the nucleus through nuclear pores.