Chapter Introduction
CHAPTER 14: Vesicular Traffic, Secretion, and Endocytosis
OUTLINE
14.1 Techniques for Studying the Secretory Pathway
14.2 Molecular Mechanisms of Vesicle Budding and Fusion
14.3 Early Stages of the Secretory Pathway
14.4 Later Stages of the Secretory Pathway
14.5 Receptor-Mediated Endocytosis
14.6 Directing Membrane Proteins and Cytosolic Materials to the Lysosome
In the previous chapter, we explored how proteins are targeted to and translocated across the membranes of several different intracellular organelles, including the endoplasmic reticulum, mitochondria, chloroplasts, peroxisomes, and the nucleus. In this chapter, we turn our attention to the secretory pathway and the mechanisms of vesicular traffic that allow proteins to be secreted from the cell or delivered to the plasma membrane and the lysosomes. We also discuss the related processes of endocytosis and autophagy, which deliver proteins and small molecules either from outside the cell or from the cytoplasm to the interior of the lysosome for degradation.
The secretory pathway is so named because it was initially studied in dedicated secretory cells that produce and secrete large quantities of proteins such as insulin or digestive enzymes to the outside of the cell. It was later discovered that the same pathway used for extracellular secretion of proteins is used to distribute all soluble and membrane proteins that enter the ER to their final destinations at the cell surface or in the lysosomes. Proteins delivered to the plasma membrane include cell-
Page 632
In contrast to the secretory pathway, which allows proteins to be targeted to the cell surface, the endocytic pathway is used to take up substances from the cell surface and move them into the interior of the cell. The endocytic pathway can selectively remove proteins from the plasma membrane and thus has a part in regulating the protein composition of the plasma membrane. In addition, the endocytic pathway is used to ingest certain nutrients that are too large to be transported across the plasma membrane by one of the transport mechanisms discussed in Chapter 11. For example, the endocytic pathway is used in the uptake of cholesterol carried in LDL particles and of iron atoms carried by the iron-
A single unifying principle governs all protein trafficking in the secretory and endocytic pathways: transport of membrane and soluble proteins from one membrane-
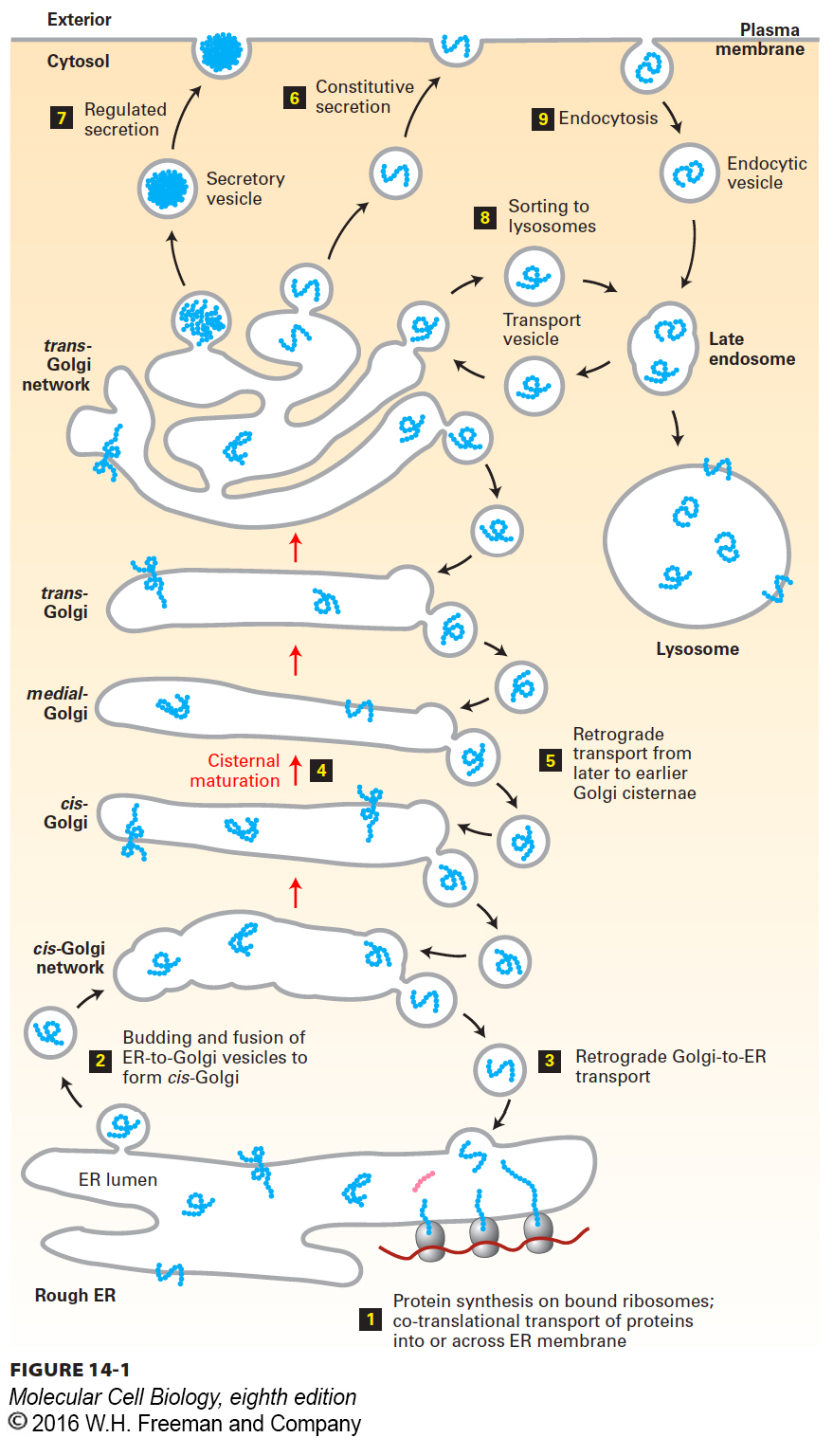
Reduced to its simplest elements, the secretory pathway operates in two stages. The first stage takes place in the rough endoplasmic reticulum (ER) (Figure 14-1, step 1). As described in Chapter 13, newly synthesized soluble and membrane proteins are translocated into the ER, where they fold into their proper conformation and receive covalent modifications such as N-linked and O-linked carbohydrates and disulfide bonds. Once newly synthesized proteins are properly folded and have received their correct modifications in the ER lumen, they progress to the second stage of the secretory pathway: transport to and through the Golgi complex.
The second stage of the secretory pathway can be summarized as follows. In the ER, cargo proteins are packaged into anterograde (forward-
Proteins in the secretory pathway are eventually delivered to a complex network of membranes and vesicles termed the trans-Golgi network. The trans-Golgi network is a major branch point in the secretory pathway. It is at this stage that proteins are loaded into different kinds of vesicles and thereby trafficked to different destinations. Depending on which kind of vesicle the protein is loaded into, it will be transported to the plasma membrane and secreted immediately, stored for later release, or shipped to the lysosome (steps 6–8). The process by which a vesicle moves to and fuses with the plasma membrane and releases its contents is known as exocytosis. In all cell types, at least some proteins are secreted continuously (a process commonly called constitutive secretion), while others are stored inside the cell until a signal for exocytosis causes them to be released (regulated secretion). Secretory proteins destined for lysosomes are first transported by vesicles from the trans-Golgi network to a compartment usually called the late endosome; the proteins are then transferred to the lysosome by direct fusion of the late endosome with the lysosomal membrane.
Endocytosis is related mechanistically to the secretory pathway. In the endocytic pathway, vesicles bud inward from the plasma membrane, bringing membrane proteins and their bound ligands into the cell (see Figure 14-1, right). After being internalized by endocytosis, some proteins are transported to lysosomes via the late endosome, whereas others are recycled back to the cell surface.
Page 633
In this chapter, we first discuss the experimental techniques that have contributed to our knowledge of the secretory pathway and endocytosis. Then we focus on the general mechanisms of membrane budding and fusion. We will see that although different kinds of transport vesicles use distinct sets of proteins for their formation and fusion, all vesicles use the same general mechanism for budding, selection of particular sets of cargo molecules, and fusion with the appropriate target membrane. In the remaining sections of the chapter, we discuss both the early and late stages of the secretory pathway, including how specificity of targeting to different destinations is achieved, and conclude with a discussion of how proteins are transported to the lysosome by the endocytic pathway.
Page 634