The 5′ Cap Is Added to Nascent RNAs Shortly After Transcription Initiation
As a nascent eukaryotic RNA transcript emerges from the RNA exit channel of RNA polymerase II (see Figure 9-12) and reaches a length of about 25 nucleotides, a protective cap composed of 7-methylguanosine and methylated riboses is added to the 5′ end of the mRNA (see Figure 5-14). This 5′ cap marks RNA molecules as mRNA precursors and protects them from RNA-digesting enzymes (5′-exoribonucleases) in the nucleus and cytoplasm. This initial step in RNA processing is catalyzed by a dimeric capping enzyme, which associates with the phosphorylated carboxy-terminal domain (CTD) of RNA polymerase II. Recall that the TFIIH general transcription factor phosphorylates the CTD multiple times on serine 5 of the CTD heptapeptide repeat during transcription initiation (see Figure 19-20). Binding of the capping enzyme to the serine 5–phosphorylated CTD stimulates the activity of the enzyme so that it is focused on RNAs containing a 5′ triphosphate that emerge from RNA polymerase II, and not on RNAs transcribed by RNA polymerases I or III, which do not have a CTD. This is important because pre-mRNA synthesis accounts for only about 80 percent of the total RNA synthesized in replicating cells. About 20 percent is pre-ribosomal RNA, which is transcribed by RNA polymerase I, and 5S rRNA, tRNAs, and other small stable RNAs, which are transcribed by RNA polymerase III. These two mechanisms, (1) binding of the capping enzyme to RNA polymerase II specifically through its unique CTD phosphorylated on serine 5 of the heptapeptide repeat during transcription initiation by TFIIH, and (2) activation of the capping enzyme by the serine 5–phosphorylated CTD, result in specific capping of RNAs transcribed by RNA polymerase II.
Page 421
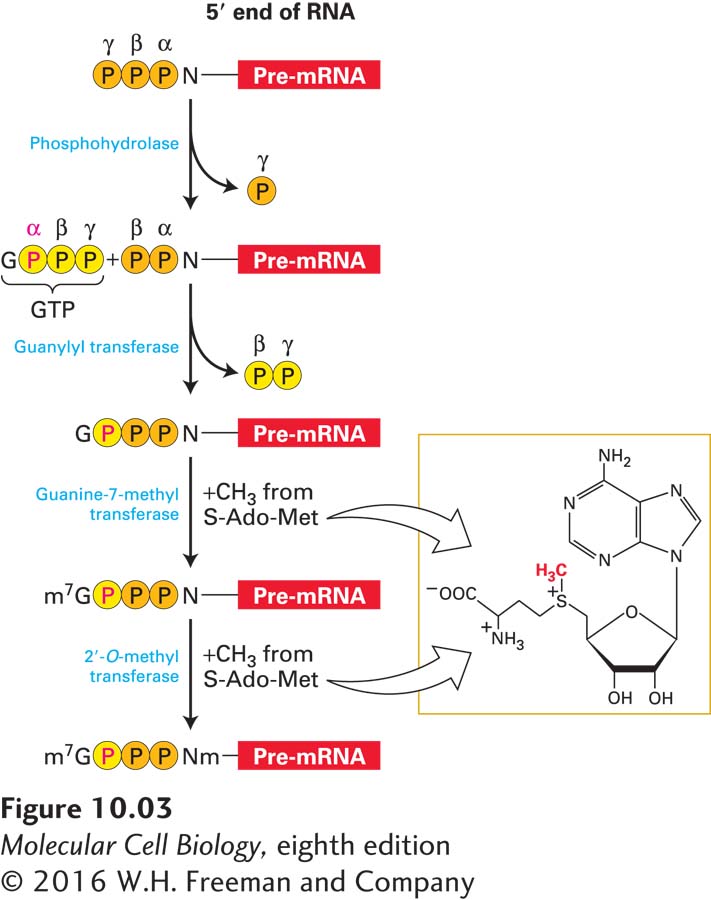
One subunit of the capping enzyme removes the γ phosphate from the 5′ end of the nascent RNA (Figure 10-3). Another domain of this subunit transfers the GMP moiety from GTP to the 5′ diphosphate of the nascent transcript, creating the unusual guanosine 5′-5′ triphosphate structure. In the final steps, separate enzymes transfer methyl groups from S-adenosylmethionine to the N7 position of the guanine and to the 2′ oxygens of riboses of the first one or two nucleotides at the 5′ end of the nascent RNA.
FIGURE 10-3Synthesis of the 5′ cap on eukaryotic mRNAs. The 5′ end of a nascent RNA contains a 5′ triphosphate from the initiating rNTP. The γ phosphate is removed in the first step of capping, while the remaining α and β phosphates (orange) remain associated with the cap. The third phosphate of the 5′,5′ triphosphate bond is derived from the α phosphate of the GTP that donates the guanine. The methyl donor for methylation of the cap guanine and the first one or two riboses of the mRNA is S-adenosylmethionine (S-Ado-Met). See S. Venkatesan and B. Moss, 1982, Proc. Natl. Acad. Sci. USA79:340.
Considerable evidence indicates that capping of the nascent transcript is coupled to elongation by RNA polymerase II so that all of its transcripts are capped during the earliest phase of elongation. As discussed in Chapter 9, in metazoans, during the initial phase of transcription, the polymerase elongates the nascent transcript very slowly due to the association of NELF (negative elongation factor) with RNA polymerase II in the promoter-proximal region (see Figure 9-21). Once the 5′ end of the nascent RNA is capped, phosphorylation of the RNA polymerase CTD at serine 2 in the heptapeptide repeat and of NELF and DSIF (DRB-sensitivity-inducing factor) by the cyclin T–CDK9 protein kinase (also known as P-TEFb) causes the release of NELF. (DRB is an analog of ATP that inhibits CDK9, preventing transcription elongation from the promoter-proximal region.) This allows RNA polymerase II to enter into a faster mode of elongation that rapidly transcribes away from the promoter. The net effect of this mechanism is that the polymerase waits for the nascent RNA to be capped before elongating at a rapid rate.