CHAPTER9:Transcriptional Control of Gene Expression
[Reproduced with permission of The Company of Biologists, from Srinivasan, S., et al., “The Drosophila trithorax group protein Kismet facilitates an early step in transcriptional elongation by RNA Polymerase II,” Development, 2005, 132(7):1623-1635; permission conveyed through Copyright Clearance Center, Inc.]
Drosophila polytene chromosomes stained with antibodies against a chromatin-remodeling ATPase called Kismet (blue), RNA polymerase II with low CTD phosphorylation (red), and RNA polymerase II with high CTD phosphorylation (green).
[Reproduced with permission of The Company of Biologists, from Srinivasan, S., et al., “The Drosophila trithorax group protein Kismet facilitates an early step in transcriptional elongation by RNA Polymerase II,” Development, 2005, 132(7):1623-1635; permission conveyed through Copyright Clearance Center, Inc.]
In previous chapters, we have seen that the properties and functions of each cell type are determined by the proteins it contains. In this chapter and the next, we consider how the kinds and amounts of the various proteins produced by a particular cell type in a multicellular organism are regulated. This regulation of gene expression is the fundamental process that controls the development of multicellular organisms such as ourselves from a single fertilized egg cell into the thousands of cell types of which we are made. When gene expression goes awry, cellular properties are altered, a process that all too often leads to the development of cancer. As discussed further in Chapter 24, genes encoding proteins that restrain cell growth are abnormally repressed in cancer cells, whereas genes encoding proteins that promote cell growth and replication are inappropriately activated in cancer cells. Abnormalities in gene expression also result in developmental defects such as cleft palate, tetralogy of Fallot (a serious developmental defect of the heart that can be treated surgically), and many others. Regulation of gene expression also plays a vital role in bacteria and other single-celled microorganisms, in which it allows cells to adjust their enzymatic machinery and structural components in response to their changing nutritional and physical environment. Consequently, to understand how microorganisms respond to their environment and how multicellular organisms normally develop, as well as how pathological abnormalities of gene expression occur, it is essential to understand the molecular interactions that control protein production.
The basic steps in gene expression—that is, the entire process whereby the information encoded in a particular gene is decoded into a particular protein—are reviewed in Chapter 5. Synthesis of mRNA requires that an RNA polymerase initiate transcription (initiation), polymerize ribonucleoside triphosphates complementary to the DNA coding strand (elongation), and then terminate transcription (termination) (see Figure 5-11). In bacteria, ribosomes and translation initiation factors have immediate access to newly formed RNA transcripts, which function as mRNA without further modification. In eukaryotes, however, the initial RNA transcript is subjected to processing that yields a functional mRNA (see Figure 5-15). The mRNA then is transported from its site of synthesis in the nucleus to the cytoplasm, where it is translated into protein with the aid of ribosomes, tRNAs, and translation factors (see Figures 5-23, 5-24, and 5-26).
Page 354
[Data from J. J. Li and M. D. Biggin, 2014, Science347:1066.]
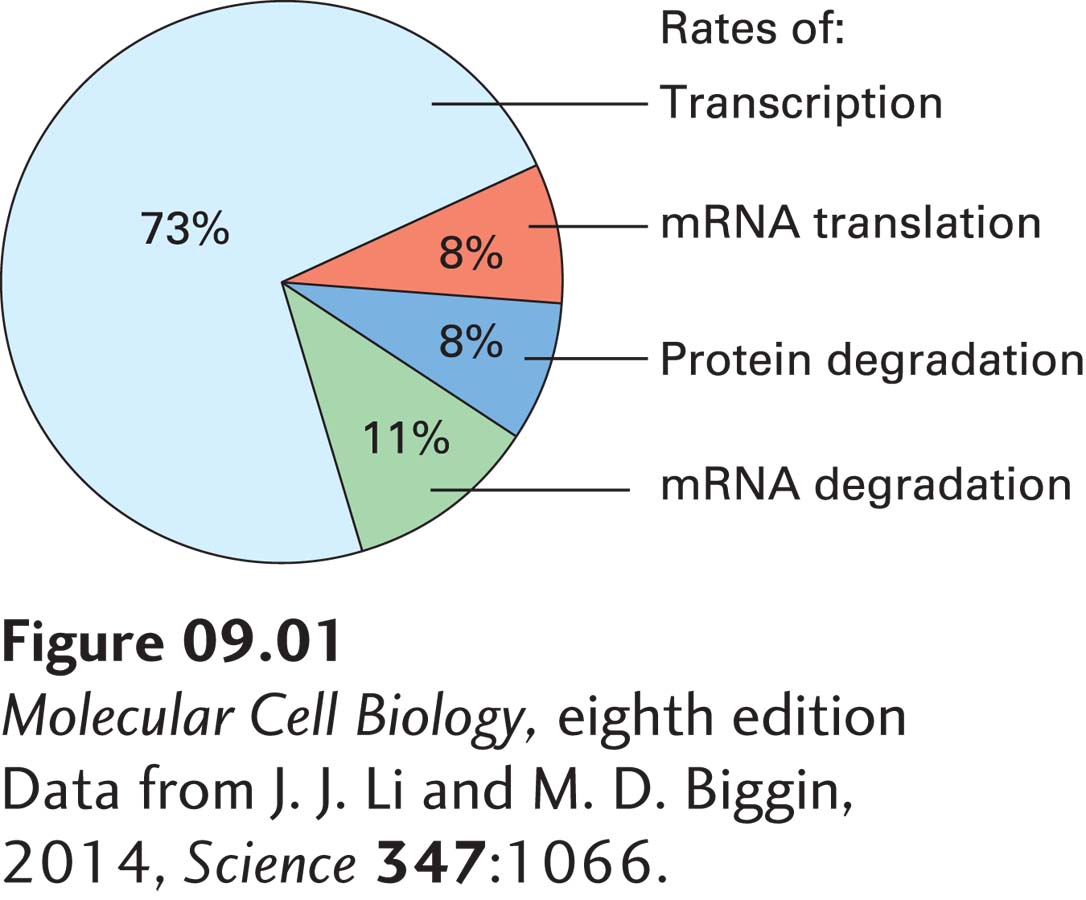
FIGURE 9-1Contributions of the major processes that regulate protein concentrations. The concentration of a protein is controlled by regulation of the frequency with which the mRNA encoding the protein is synthesized (gene transcription), the rate at which that mRNA is degraded, the rate at which that mRNA is translated into protein, and the rate at which that protein is degraded. The relative contributions of these four rates to determining the concentrations of thousands of proteins in cultured mouse fibroblasts were determined by mass spectrometry to measure protein concentrations (see Chapter 3), mRNA sequencing (RNA-seq) to measure mRNA levels (see Chapter 6), protection of mRNA from ribonuclease digestion by associated ribosomes (ribosome footprinting) to estimate translation rates, stable isotope labeling to determine degradation rates, and statistical analysis of the data to correct for inherent biases and errors in these methods.
[Data from J. J. Li and M. D. Biggin, 2014, Science347:1066.]
Regulation may occur at several of the various steps in gene expression outlined above: transcription initiation, elongation, RNA processing, and mRNA export from the nucleus, as well as through control of mRNA degradation, mRNA translation into protein, and protein degradation. This regulation results in differential protein expression in different cell types or developmental stages or in response to external conditions. Although examples of regulation at each step in gene expression have been found, control of transcription initiation and of elongation—the first two steps—are the most important mechanisms for determining whether most genes are expressed and how much of the encoded mRNAs and, consequently, proteins are produced (Figure 9-1). The molecular mechanisms that regulate transcription initiation and elongation are critical to numerous biological phenomena, including the development of a multicellular organism, as mentioned above, the immune responses that protect us from pathogenic microorganisms, and neurological processes such as learning and memory. When these regulatory mechanisms controlling transcription function improperly, pathological processes may occur. For example, dominant mutations of the HOXD13 gene result in polydactyly, the embryological development of extra digits of the feet, hands, or both (Figure 9-2a). HOXD13 encodes a transcription factor that normally regulates the transcription of multiple genes involved in development of the extremities. Other mutations affecting the function or expression of transcription factors cause an extra pair of wings to develop in Drosophila (Figure 9-2b), alter the structures of flowers in plants (Figure 9-2c), and are responsible for multiple other developmental abnormalities.
[Part (a), left, Lightvision, LLC/Moment Open/Getty Images; right, Goodman, F. R. and Scrambler, P. J., Human HOX gene mutations. Clinical Genetics, 2001, 59:1, pages 1–11. Part (b) from “The bithorax complex: the first fifty years,” by Edward B. Lewis, reproduced with permission from The International Journal of Developmental Biology, 1998, Vol 42(403-15), Figures 4a and 4b. Part (c) republished with permission of Elsevier, from Weigel, D. and Meyerowitz, M., “The ABCs of floral homeotic genes,” Cell, 1994, 78(2):203-209; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 9-2Phenotypes of mutations in genes encoding transcription factors. (a) A dominant mutation in the human HOXD13 gene results in the development of extra digits, a condition known as polydactyly. (b) Homozygous recessive mutations that prevent expression of the Ubx gene in the third thoracic segment of Drosophila result in transformation of that segment, which normally has a balancing organ called a haltere, into a second copy of the thoracic segment that develops wings. (c) Mutations in Arabidopsis thaliana that inactivate both copies of three floral organ–identity genes transform the normal parts of the flower into leaflike structures. In each case, these mutations affect master regulatory transcription factors that regulate multiple genes, including many genes encoding other transcription factors.
[Part (a), left, Lightvision, LLC/Moment Open/Getty Images; right, Goodman, F. R. and Scrambler, P. J., Human HOX gene mutations. Clinical Genetics, 2001, 59:1, pages 1–11. Part (b) from “The bithorax complex: the first fifty years,” by Edward B. Lewis, reproduced with permission from The International Journal of Developmental Biology, 1998, Vol 42(403-15), Figures 4a and 4b. Part (c) republished with permission of Elsevier, from Weigel, D. and Meyerowitz, M., “The ABCs of floral homeotic genes,” Cell, 1994, 78(2):203-209; permission conveyed through Copyright Clearance Center, Inc.]
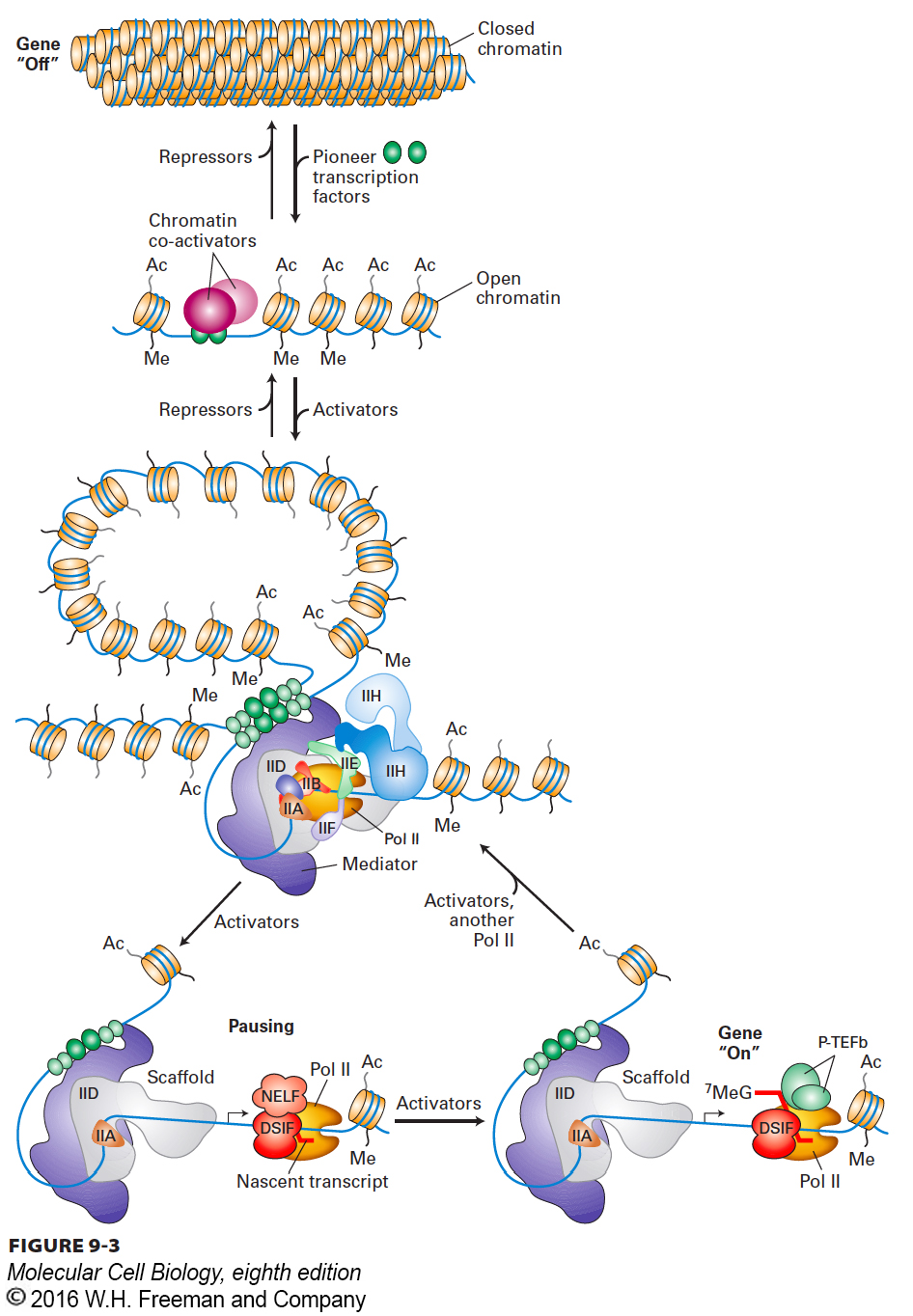
Transcription is a complex process involving many layers of regulation. In this chapter, we focus on the molecular events that determine when transcription of a gene occurs. First, we consider the mechanisms of gene expression in bacteria, in which DNA is not bound by histones and packaged into nucleosomes. Repressor and activator proteins recognize and bind to specific DNA sequences to control the transcription of a nearby gene, and in many cases, specific tertiary structures in nascent mRNAs, called riboswitches, bind metabolites to regulate transcription elongation. The remainder of the chapter focuses on eukaryotic regulation of transcription and how the basic tenets of bacterial regulation are applied in more complex ways in higher organisms. In addition, eukaryotic regulation mechanisms make use of the association of DNA with histone octamers, forming chromatin structures with varying degrees of condensation, and of post-translational modifications of histone tails such as acetylation and methylation (see Figure 8-26). Figure 9-3 provides an overview of transcriptional regulation in metazoans (multicellular animals) and of the processes outlined in this chapter. We discuss how the RNA polymerases responsible for the transcription of different classes of eukaryotic genes bind to promoter sequences to initiate the synthesis of an RNA molecule, and how specific DNA sequences function as transcription-control regions by serving as the binding sites for the transcription factors that regulate transcription. Next we consider how eukaryotic activators and repressors influence transcription through interactions with large multiprotein complexes. Some of these multiprotein complexes modify chromatin condensation, altering the accessibility of chromosomal DNA to transcription factors and RNA polymerases. Other complexes directly influence the frequency at which RNA polymerases bind to promoters and initiate transcription. Very recent research has revealed that, for many genes in multicellular animals, the RNA polymerase pauses after transcribing a short RNA, and that one transcriptional regulation mechanism involves a release of the paused polymerase, allowing it to transcribe the rest of the gene. We discuss how transcription of specific genes can be specified by particular combinations of the roughly 1400 transcription factors encoded in the human genome, giving rise to cell-type-specific gene expression. We consider the various ways in which the activities of transcription factors themselves are controlled to ensure that genes are expressed only in the correct cell types and at the appropriate time during their differentiation.
FIGURE 9-3Overview of eukaryotic transcriptional control. Inactive genes are assembled into regions of condensed chromatin that inhibit RNA polymerases and their associated general transcription factors from interacting with promoters. A pioneer transcription factor is able to bind to a specific regulatory sequence within the condensed chromatin and interact with chromatin-remodeling enzymes and histone acetylases that decondense the chromatin, making it accessible to RNA polymerase II and the general transcription factors. Additional activator proteins then bind to specific transcription-control elements in both promoter-proximal sites and distant enhancers, where they interact with one another and with the multisubunit Mediator complex to assemble RNA polymerase II (Pol II) and general transcription factors on promoters. Alternatively, repressor proteins bind to other transcription-control elements to inhibit transcription initiation by Pol II and interact with multiprotein co-repressor complexes to condense chromatin. During transcriptional activation, Pol II initiates transcription, but pauses after transcribing fewer than 100 nucleotides due to the action of the elongation inhibitor NELF associated with DSIF. Activators promote the association of the Pol II-NELF-DSIF complex with elongation factor P-TEFb, which releases NELF and allows productive elongation through the gene. DSIF is the DRB sensitivity-inducing factor, NELF is the negative elongation factor, and P-TEFb is a protein kinase made up of CDK9 and cyclin T. See S. Malik and R. G. Roeder, 2010, Nat. Rev. Genet.11:761.
We also discuss recent studies revealing that RNA-protein complexes in the nucleus can regulate transcription. New methods for sequencing DNA, coupled with reverse transcription of RNA into DNA in vitro, have revealed that much of the genome of eukaryotes is transcribed into low-abundance RNAs that do not encode proteins. Several nuclear long noncoding RNAs (lncRNAs) have recently been discovered to regulate the transcription of other protein-coding genes. This finding raises the possibility that transcriptional control by such noncoding RNAs may be much more general than is currently understood. Recent advances in mapping the association of transcription factors with specific regions of chromatin across the entire genome in a variety of cell types have provided the first glimpses of how transcription factors regulate embryonic development from the pluripotent stem cells of the early embryo to the fully differentiated cells that make up most of our tissues. RNA processing and various post-transcriptional mechanisms for controlling eukaryotic gene expression are covered in Chapter 10. Subsequent chapters, particularly Chapters 15, 16, and 21, provide examples of how transcription is regulated by interactions between cells and how the resulting gene control contributes to the development and function of specific types of cells in multicellular organisms.