CHAPTER15:Signal Transduction and G Protein–Coupled Receptors
[Data from Y. Kang et al., 2015, Nature523:561-567, PDB ID 4zwj, and custom PDB.]
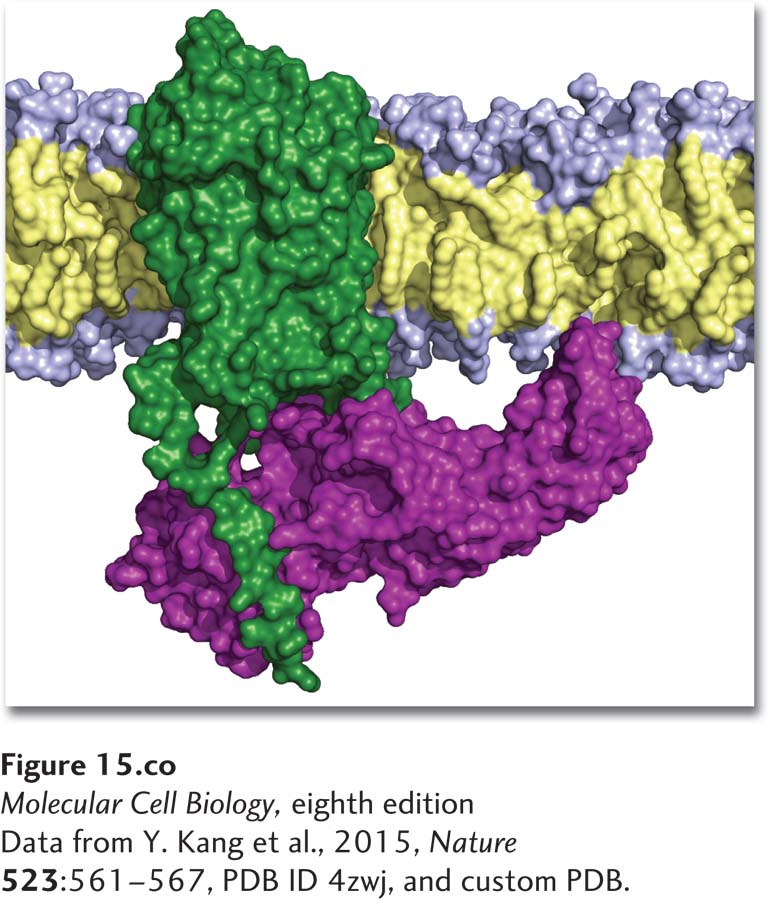
Structure of a cell surface G protein–coupled receptor (green) bound to β-arrestin (purple). G protein–coupled receptors that are in the active state for a long period of time become phosphorylated, triggering binding of an arrestin and inhibition of further signaling by the receptor.
[Data from Y. Kang et al., 2015, Nature523:561-567, PDB ID 4zwj, and custom PDB.]
No cell lives in isolation; life requires that all cells sense chemicals and physical stimuli in their environment and respond with changes that can affect their function or development. Many cells sense and respond to light: many single-celled algae are phototactic (see Chapter 1), and later in this chapter, we will see how cells in the human retina sense light and respond by sending signals to the brain. Other cells sense and respond to physical stimuli such as touch or heat.
Most signals are chemical molecules; many, such as odorants, tastants (substances that can be tasted), and even oxygen are in the environment of virtually every metazoan cell and organism. Many types of signaling molecules are released by one cell and induce a response in a different cell; this fundamental process, known as cellular communication, shapes the development and function of every living organism. Even single-celled eukaryotic microorganisms, such as yeasts, algae, slime molds, and protozoans, communicate through extracellular signals. In the next chapter, we will see how secreted molecules called pheromones coordinate the aggregation of free-living yeast cells for sexual mating.
More important in multicellular plants and animals are hormones and other extracellular signaling molecules that function within an organism to control a variety of processes, including the metabolism of sugars, fats, and amino acids; the growth and differentiation of tissues; the synthesis and secretion of small-molecule hormones and many proteins; and the composition of intracellular and extracellular fluids. Many types of metazoan cells also respond to signals from the external environment, including light, oxygen, odorants, and tastants in food.
In any system, for a signal to have an effect on a target, it has to be received. In cells, a signal produces a specific response only in target cells with receptor proteins that bind that signal. For some receptors, this signal is light, touch, or heat. Many types of chemicals act as signals: small molecules (e.g., amino acid and lipid derivatives, steroids, acetylcholine), gases (e.g., oxygen, nitric oxide), peptides (e.g., adrenocorticotropic hormone and vasopressin), soluble proteins (e.g., insulin and growth hormone), and proteins that are tethered to the surface of a cell or bound to the extracellular matrix. Many of these extracellular signaling molecules are synthesized, packaged into secretory vesicles, and released by specialized signaling cells within multicellular organisms. Like enzymes, receptors bind a single type of molecule or a group of closely related molecules; unlike enzymes, receptors do not catalyze a chemical transformation of the bound molecule.
Page 674
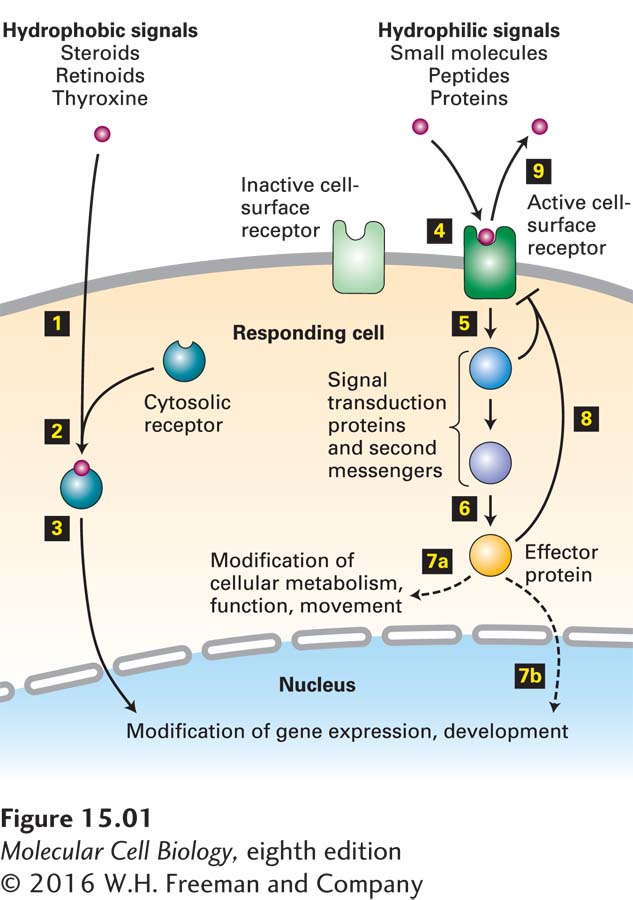
One large class of hydrophobic signaling molecules, primarily molecules such as steroids, retinoids, and thyroxine, spontaneously diffuse through the plasma membrane and bind to receptors in the cytosol (Figure 15-1). In most cases, as we saw in Chapter 9, the receptor-hormone complex moves into the nucleus, binds to specific regulatory sequences in DNA, and activates or represses expression of specific target genes.
FIGURE 15-1Overview of cell signaling. Hydrophobic signaling molecules, such as steroids and related molecules, diffuse through the plasma membrane (step 1) and bind to receptors in the cytosol (step 2). The receptor-signal complex moves into the nucleus (step 3), where it can bind to transcription-control regions in DNA and activate or repress gene expression. The majority of signaling molecules, including small molecules (adrenaline, acetylcholine), peptides (yeast mating factors, glucagon), and proteins (insulin, growth hormone), are hydrophilic and cannot diffuse across the cell membrane. These molecules bind to specific cell-surface receptor proteins, triggering a conformational change in the receptor, thus activating it (step 4). The activated receptor then activates one or more downstream signal transduction proteins or small-molecule second messengers (step 5), which eventually leads to activation of one or more effector proteins (step 6). The end result of a signaling cascade can be modification of specific cytosolic proteins, predominantly enzymes, leading to short-term changes in cellular function, metabolism, or movement (step 7a). Alternatively, an effector can move into the nucleus, triggering a long-term change in gene expression (step 7b). Termination or down-modulation of the cellular response is often caused by negative feedback from intracellular signaling molecules (step 8) and by removal of the extracellular signal (step 9).
In this chapter and the next, we focus on extracellular signaling molecules that are too large and too hydrophilic to diffuse through the plasma membrane. How, then, can they affect intracellular processes? These signaling molecules bind to cell-surface receptors that are integral membrane proteins embedded in the plasma membrane. Cell-surface receptors generally consist of three discrete topological domains, or segments: an extracellular domain facing the extracellular fluid, a plasma-membrane-spanning (transmembrane) domain, and an intracellular domain facing the cytosol. The signaling molecule acts as a ligand, which binds to a structurally complementary site on the extracellular or the membrane-spanning domain of the receptor. Binding of the ligand to its site on the receptor induces a conformational change in the receptor that is transmitted through the membrane-spanning domain to the cytosolic domain. This conformational change can result in the receptor’s binding to, and subsequent activation or inhibition of, other proteins in the cytosol or attached to the plasma membrane. In many cases, these activated proteins catalyze the synthesis of specific small molecules or change the concentration of an intracellular ion such as Ca2+. These intracellular small-molecule second messengers then carry the signal to one or more effector proteins, such as enzymes or transcription factors.
Many signaling proteins, including the GTP-binding “switch” proteins and protein kinases introduced in Chapter 3, are members of large classes of signal transduction proteins that have been highly conserved throughout evolution. The overall process of converting extracellular signals into intracellular responses, as well as the individual steps in this process, is termed signal transduction; the chain of intermediates is called a signal transduction pathway because it transduces, or converts, information from one form into another as a signal is relayed from a receptor to its targets. Some signal transduction pathways contain just two or three intermediates; others can involve over a dozen.
As shown in Figure 15-1, signal transduction, which begins when extracellular signaling molecules bind to cell-surface receptors, can induce two major types of cellular responses. The first (step 7a) involves short-term (seconds to minutes) changes in the activity or function of specific enzymes and other proteins that preexist in the cell, often by covalent modifications such as phosphorylation or ubiquitinylation or by binding of molecules such as cAMP or Ca2+. The second (step 7b) involves activation (or repression) of specific transcription factors, often by phosphorylation or other covalent modifications, leading to long-term (hours to days) changes in the amounts of specific proteins contained in a cell.
Page 675
In eukaryotes, there are about a dozen classes of cell-surface receptors, which activate several types of intracellular signal transduction pathways. Our knowledge of signal transduction has advanced greatly in recent years, in large measure because these receptors and pathways are highly conserved and function in essentially the same way in organisms as diverse as worms, flies, mice, and humans. Genetic studies combined with biochemical analyses have enabled researchers to trace many entire signaling pathways from binding of ligand to final cellular responses. In this chapter, we first review some general principles of signal transduction, such as the molecular basis for ligand-receptor binding, and certain evolutionarily conserved components of signal transduction pathways. Next we describe how cell-surface receptors and signal transduction proteins are identified and characterized biochemically.
We then turn to an in-depth discussion of a very large and evolutionarily conserved class of receptors, found in organisms from fungi to humans: the G protein–coupled receptors. As their name implies, G protein–coupled receptors (GPCRs) consist of an integral membrane receptor protein coupled to an intracellular G protein that transmits signals to the interior of the cell. The human genome encodes more than 800 G protein–coupled receptors, which constitute about 4 percent of the identified human proteins. Most bind specific small molecules or peptides, but a few bind proteins. GPCRs include receptors in the visual, olfactory (smell), and gustatory (taste) systems, many neurotransmitter receptors, and most of the receptors for hormones that control carbohydrate, amino acid, and fat metabolism and even behavior. Signal transduction through GPCRs initially induces short-term changes in cell function, such as a change in metabolism or movement, but many of these signal transduction pathways also lead to changes in gene expression. We show how these pathways affect many aspects of cell function, including glucose metabolism, muscle contraction, perception of light, and gene expression.
Other large classes of cell-surface receptors mainly bind protein ligands. Activation of these receptors primarily alters a cell’s pattern of gene expression, leading to cell differentiation or division and other long-term consequences, but can also induce short-term effects on cells. These receptors and the intracellular signaling pathways they activate are explored in Chapter 16. In Chapter 22, we discuss other classes of receptors, mainly confined to the nervous system, that are coupled to ion channels.