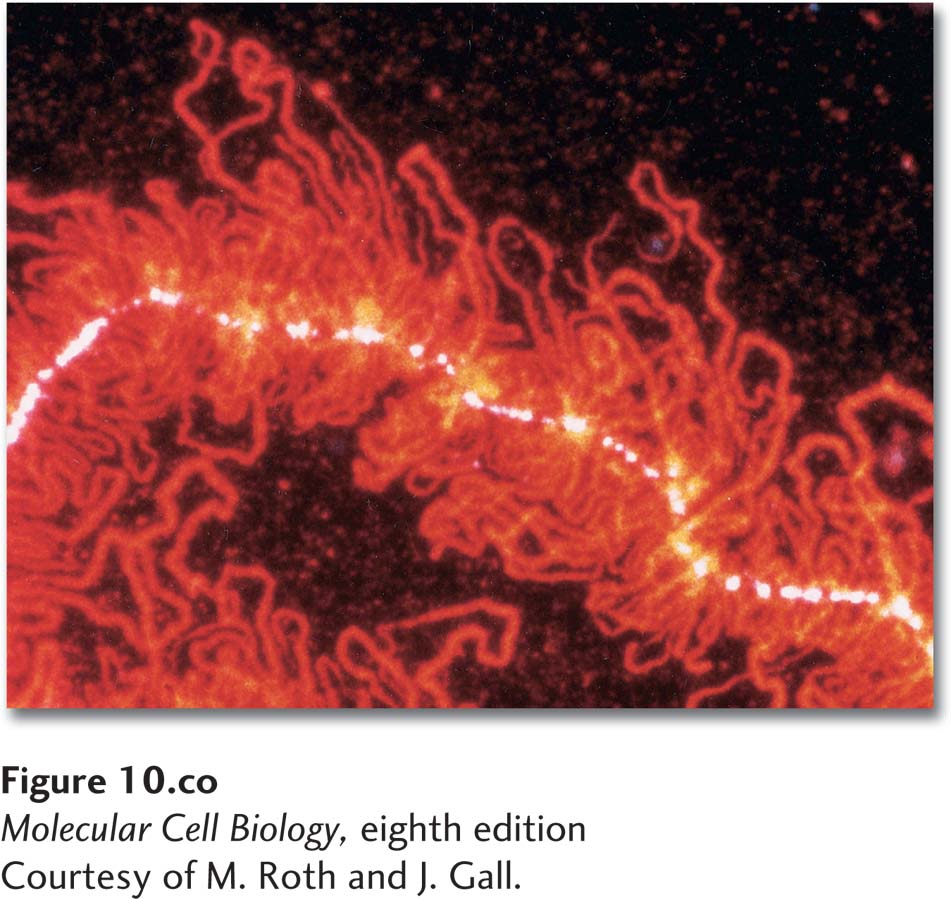
Portion of a “lampbrush chromosome” from an oocyte of the newt Nophthalmus viridescens. The hnRNP protein associated with nascent RNA transcripts fluoresces red after staining with a monoclonal antibody.
In the previous chapter, we saw that most genes are regulated at the first step in gene expression, transcription, by regulation of the assembly of the transcription preinitiation complex on a promoter DNA sequence and of transcription elongation in the promoter-proximal region. Once transcription has been initiated, synthesis of the encoded RNA requires that RNA polymerase transcribe the entire gene and not terminate transcription prematurely. Moreover, the initial primary transcripts produced from eukaryotic genes must undergo various processing reactions to yield the corresponding functional RNAs. For mRNAs, the 5′ cap structure necessary for translation must be added (see Figure 5-14), introns must be spliced out of pre-mRNAs, and the 3′ end must be polyadenylated (see Figure 5-15). Once formed in the nucleus, mature, functional RNAs are exported to the cytoplasm as components of ribonucleoproteins. Both the processing of RNAs and their export from the nucleus offer opportunities for further regulation of gene expression after the initiation of transcription.
Recently, the vast amount of sequence data on human mRNAs expressed in different tissues and at various times during embryogenesis and cellular differentiation has revealed that some 95 percent of human genes give rise to alternatively spliced mRNAs. These alternatively spliced mRNAs encode related proteins with differences in their sequences that are limited to specific functional domains. In many cases, alternative RNA splicing is regulated to meet the need for a specific protein isoform in a specific cell type. Given the complexity of pre-mRNA splicing, it is not surprising that mistakes are occasionally made, giving rise to mRNA precursors with improperly spliced exons. However, eukaryotic cells have evolved RNA surveillance mechanisms that prevent the export of incorrectly processed RNAs to the cytoplasm or lead to their degradation if they are exported.
Additional control of gene expression can occur in the cytoplasm. In the case of protein-coding genes, for instance, the amount of protein produced depends on the stability of the corresponding mRNAs in the cytoplasm and the rate of their translation. For example, during an immune response, lymphocytes communicate by secreting polypeptide hormones called cytokines that signal neighboring lymphocytes through cytokine receptors that span their plasma membranes (see Chapter 23). It is important for lymphocytes to synthesize and secrete cytokines in short bursts. This is possible because cytokine mRNAs are extremely unstable; consequently, the concentration of these mRNAs in the cytoplasm falls rapidly once their synthesis is stopped. In contrast, mRNAs encoding proteins required in large amounts that function over long periods, such as ribosomal proteins, are extremely stable, so that multiple polypeptides are transcribed from each mRNA.
Page 418
Page 419
Just as pre-mRNA processing, nuclear export, and translation are regulated, so is the cellular localization of many, if not most, mRNAs, so that newly synthesized protein is concentrated where it is needed. Particularly striking examples of this type of regulation occur in the nervous systems of multicellular animals. Some neurons in the human brain generate more than a thousand separate synapses with other neurons. During the process of learning, synapses that fire more frequently than others increase in size many times, while other synapses made by the same neuron do not. This can occur because mRNAs encoding proteins critical for synapse enlargement are stored at all synapses, but translation of these localized, stored mRNAs is regulated at each synapse independently by the frequency at which the synapse signals. In this way, synthesis of synapse-associated proteins can be regulated independently at each of the many synapses made by the same neuron (see Chapter 22).
Another type of gene regulation involves micro-RNAs (miRNAs), which regulate the translation and stability of specific target mRNAs in multicellular animals and plants. Analyses of these short miRNAs in various human tissues indicate that about 1900 miRNAs are expressed in the multiple types of human cells. Although some have recently been discovered to function through inhibition of target-gene expression in the appropriate tissue and at the appropriate time in development, the functions of the vast majority of human miRNAs are unknown and are the subject of a growing new area of research. If most miRNAs do indeed have significant functions, miRNA genes constitute an important subset of the 25,000 or so human genes. A closely related process, called RNA interference (RNAi), leads to the degradation of viral RNAs in infected cells and the degradation of transposon-encoded RNAs in many eukaryotes. This discovery is of tremendous significance to biological researchers because it is possible to design short interfering RNAs (siRNAs) to inhibit the translation of specific mRNAs experimentally by a process called RNA knockdown. This method makes it possible to inhibit the function of any desired gene, even in organisms that are not amenable to classical genetic methods for isolating mutants.
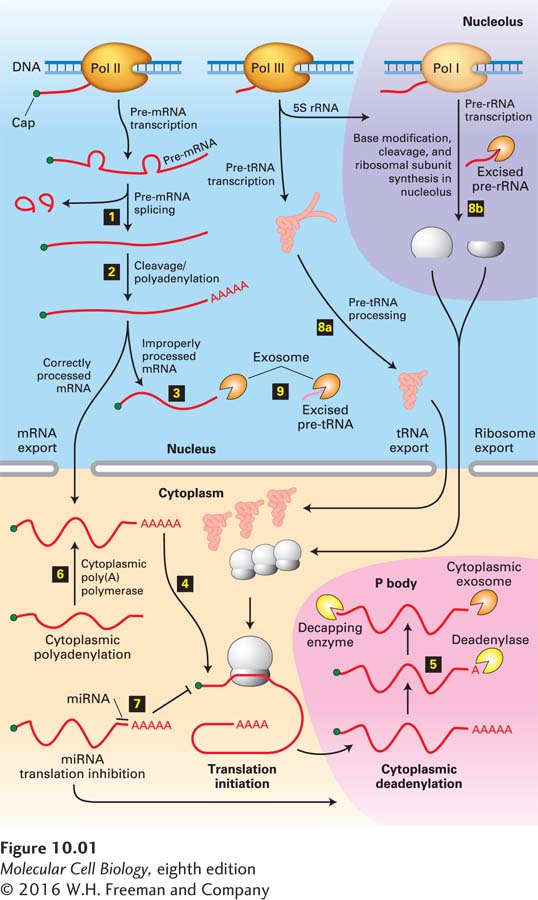
We refer to all the mechanisms that regulate gene expression following transcription as post-transcriptional gene control (Figure 10-1). Because the stability and translation rate of an mRNA contribute to the amount of protein expressed from a gene, these post-transcriptional processes are important components of gene control. Indeed, the protein output of a gene is regulated at every step in the life of an mRNA, from the initiation of its synthesis to its degradation. Thus genetic regulatory processes act on RNA as well as on DNA. In this chapter, we consider the events in the processing of mRNA that follow transcription initiation and promoter-proximal elongation as well as the various mechanisms that are known to regulate these events. In the last section, we briefly discuss the processing of primary transcripts produced from genes encoding rRNAs and tRNAs.
FIGURE 10-1Overview of RNA processing and post-transcriptional gene control. Nearly all cytoplasmic RNAs are processed from primary transcripts in the nucleus before they are exported to the cytoplasm. For protein-coding genes transcribed by RNA polymerase II, gene control can be exerted through step 1 the choice of alternative exons during pre-mRNA splicing and step 2 the choice of alternative poly(A) sites. Improperly processed mRNAs are blocked from export to the cytoplasm and degraded step 3 by a large complex called the exosome that contains multiple ribonucleases. Once the mRNA has been exported to the cytoplasm, step 4 translation initiation factors bind to the 5′ cap cooperatively with poly(A)-binding protein I bound to the poly(A) tail and initiate translation (see Figure 4-28). Step 5 mRNA is degraded in the cytoplasm by deadenylation and decapping followed by degradation by cytoplasmic exosomes. These processes occur rapidly in dense regions of the cytoplasm called P bodies that function in translational repression. The degradation rate of each mRNA is controlled, thereby regulating the mRNA concentration and, consequently, the amount of protein translated. Some mRNAs are synthesized without long poly(A) tails. Their translation is regulated by step 6 control of the synthesis of a long poly(A) tail by a cytoplasmic poly(A) polymerase. Step 7 Translation is also regulated by other mechanisms, including miRNAs. When expressed, these ~22-nucleotide RNAs inhibit translation of mRNAs to which they hybridize, usually in the 3′ untranslated region. tRNAs and rRNAs are also synthesized as precursor RNAs that must be step 8 processed before they are functional. Regions of precursors cleaved from the mature RNAs are degraded by nuclear exosomes step 9. See Houseley et al., 2006, Nat. Rev. Mol. Cell Biol.7:529.