Chapter Introduction
CHAPTER 21: Stem Cells, Cell Asymmetry, and Cell Death

Many descriptions of cell division imply that the parent cell gives rise to two daughter cells that look and function exactly like the parent cell. In other words, they imply that cell division is symmetric and that the progeny have properties similar to those of the parent (Figure 21-1a). Many yeasts, fungi, and other single-
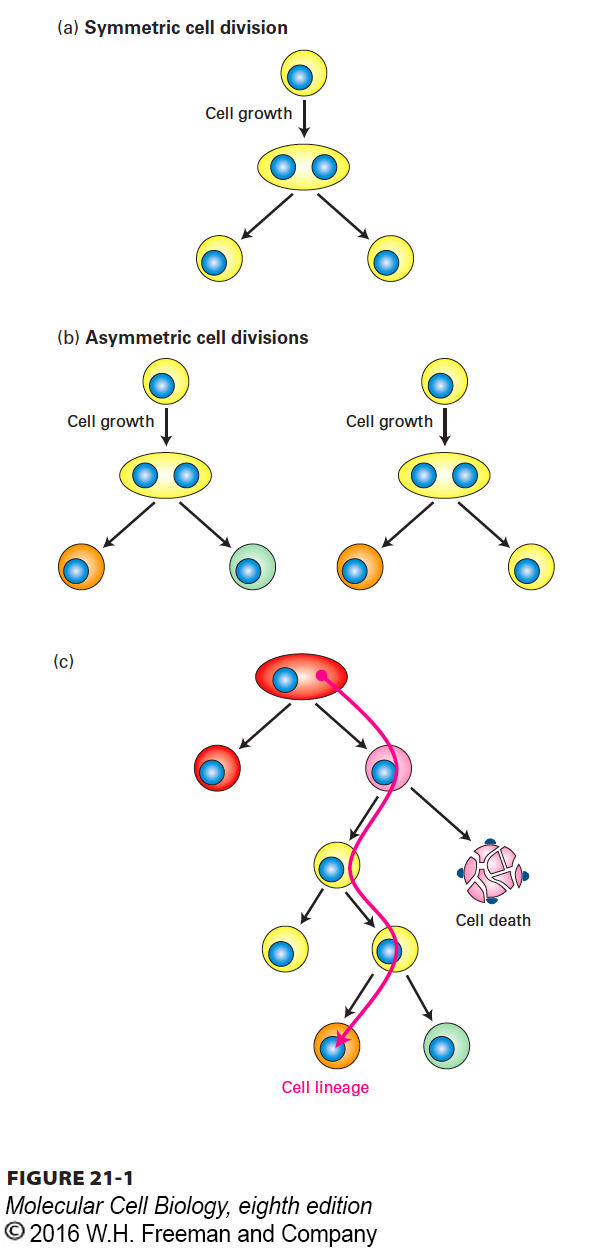
But if this were always the case, none of the hundreds of differentiated cell types and functioning tissues present in complex multicellular plants and animals would ever be formed. Differences among cells can arise when two initially identical daughter cells diverge upon receiving distinct developmental or environmental signals. Alternatively, the two daughter cells may differ from “birth,” with each inheriting different portions of the parent cell (Figure 21-1b). Daughter cells produced by such asymmetric cell division may differ in size, shape, or protein composition, or their genes may be in different states of activity or potential activity. The differences in these internal signals confer different fates on the two cells. In certain asymmetric cell divisions, one of the daughters is similar to the parent cell and the other forms a different type of cell.
In multicellular organisms, the formation of working tissues and organs, during both development and cell replacement, depends on specific patterns of mitotic cell divisions. A series of such cell divisions akin to a family tree is called a cell lineage. A cell lineage traces the birth order of cells as they progressively become more restricted in their developmental potential and differentiate into specialized cell types such as skin cells, neurons, or muscle cells (Figure 21-1c).
The development of a new metazoan organism begins with the egg, or oocyte, carrying a set of chromosomes from the mother, and the sperm, carrying a set of chromosomes from the father. These gametes, or sex cells, are haploid because they have gone through meiosis (see Chapter 19). In the process called fertilization, they combine to create the initial single cell, the zygote, which has two sets of chromosomes and is therefore diploid. During embryogenesis, the zygote undergoes numerous cell divisions, both symmetric and asymmetric, ultimately giving rise to an entire organism. As we will see later in the chapter (see Figure 21-25), many of the early divisions of the nematode Caenorhabditis elegans follow a mosaic development strategy, in which all of the early cell divisions are asymmetric and each daughter cell gives rise to a discrete set of differentiated cell types because regulatory proteins located in cytoplasmic granules are unequally distributed to the daughter cells.
Page 976
The focus of the first section of this chapter is early mammalian development and its regulation by cell-
Stem cells are important during both metazoan development and adult life. They are unspecialized cells that can reproduce themselves as well as generate specific types of more specialized cells (see Figure 21-1b). Their name comes from the image of a plant stem, which grows upward, continuing to form more stem, while also sending off leaves and branches to the side. In the second and third sections of this chapter, we explore several types of stem cells that differ in the variety of specialized cell types they can form. Stem cells can undergo symmetric divisions in which both of the daughters are stem cells. Many types of stem cells in animals and plants undergo asymmetric divisions in which one of the daughter cells is a stem cell. Thus the numbers of stem cells can remain constant or can increase during the organism’s life. The zygote is totipotent in that it has the capacity to generate every cell type in the body as well as the supportive placental cells that are required for embryonic development, but because the zygote does not self-
Page 977
In Section 21.2, we will learn that cells of the inner cell mass can be cultured in defined media, forming embryonic stem (ES) cells. ES cells can be grown indefinitely in culture, where they divide symmetrically, so that each daughter cell remains pluripotent and can potentially give rise to all of the tissues of an animal. We will discuss the use of ES cells in uncovering the transcriptional network of gene expression underlying pluripotency as well as in forming specific types of differentiated cells for research purposes or, potentially, as “replacement parts” for worn-
For many years, animal cell differentiation was thought to be unidirectional, but recent data reveal that differentiation can be reversed experimentally. Through recombinant expression of specific transcription factors, one type of specialized, differentiated cell can be converted into another type of differentiated cell. Strikingly, introducing just a small number of the transcription factors that control the pluripotency of ES cells into multiple types of differentiated cells, under defined conditions, can convert at least some of those somatic cells into induced pluripotent stem (iPS) cells that have properties seemingly indistinguishable from those of ES cells. As we will see in Section 21.2, iPS cells have profound utility for experimental biology and medicine.
Many types of cells have life spans much shorter than that of the organism as a whole and so need to be constantly replaced. In mammals, for instance, cells lining the intestine and phagocytic macrophages live for only a few days. Stem cells are therefore important not only during development, but also for replacement of worn-
We have already mentioned that the diversity of cell types in an animal requires asymmetric cell divisions in which the fates of the two daughter cells differ. This process requires the parent cell to become asymmetric, or polarized, before cell division, so that the cell contents are unequally distributed between the two daughters. This process of polarization is critical not only during development, but also for the function of essentially all cells. For example, transporting epithelial cells, such as those that line the intestine, are polarized, with their free apical surface facing the lumen to absorb nutrients and their basolateral surface contacting the extracellular matrix to transport nutrients toward the blood (see Figures 11-30 and 20-1). Other examples include cells that migrate up a chemotactic gradient (see Figure 18-53) and neurons, which have multiple dendrites extending from one side of the cell body that receive signals and a single axon extending from the other side that transmits signals to target cells (see Chapter 22). Thus the mechanisms that cells use to polarize are important and general aspects of their function. Not surprisingly, these mechanisms integrate elements of cell signaling pathways (see Chapters 15 and 16), cytoskeletal reorganization (see Chapters 17 and 18), and membrane trafficking (see Chapter 14). In the fourth section of this chapter, we discuss how cells become polarized as well as the importance of asymmetric cell division for maintaining stem cells and generating differentiated cells.
Typically, we think of cell fates in terms of the differentiated cell types that are formed. A quite different cell fate, programmed cell death, is also absolutely crucial in the formation and maintenance of many tissues. A precise genetic regulatory system, with checks and balances, controls cell death, just as other genetic programs control cell division and differentiation. In the last section of this chapter, we consider the mechanisms of cell death and their regulation.
These aspects of cell biology—