CHAPTER17:Cell Organization and Movement I: Microfilaments
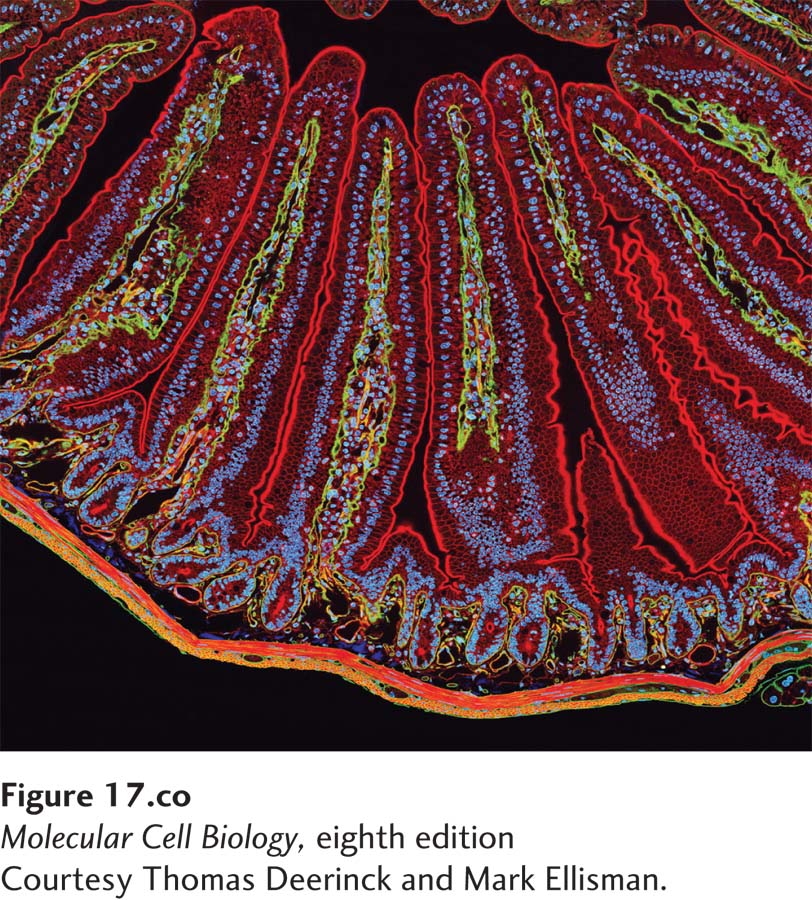
[Courtesy Thomas Deerinck and Mark Ellisman.]
A section of mouse intestine stained for actin (red), the extracellular matrix protein laminin (green), and DNA (blue). Each blue dot of DNA indicates the presence of a cell. Actin in the microvilli on the apical end of the epithelial cells can be seen lining the surface facing the lumen (top). Actin can also be seen prominently in the smooth muscle that surrounds the intestine (bottom).
When we look through a microscope at the wonderful diversity of cells in nature, the variety of cell shapes and movements we can discern is astonishing. At first we may notice that some cells, such as vertebrate sperm, ciliates such as Tetrahymena, or flagellates such as Chlamydomonas, swim rapidly, propelled by cilia and flagella. Other cells, such as amoebae and human macrophages, move more sedately, propelled not by external appendages, but by coordinated movement of the cell itself. We might also notice that some cells in tissues attach to one another, forming a pavement-like sheet, whereas other cells—neurons, for example—have long processes, up to 3 feet in length, and make selective contacts between cells. Looking more closely at the internal organization of cells, we see that organelles have characteristic locations; for example, the Golgi complex is generally near the central nucleus. How is this diversity of shape, cellular organization, and motility achieved? Why is it important for cells to have a distinct shape and clear internal organization? Let us first consider two examples of cells with very different functions and organizations.
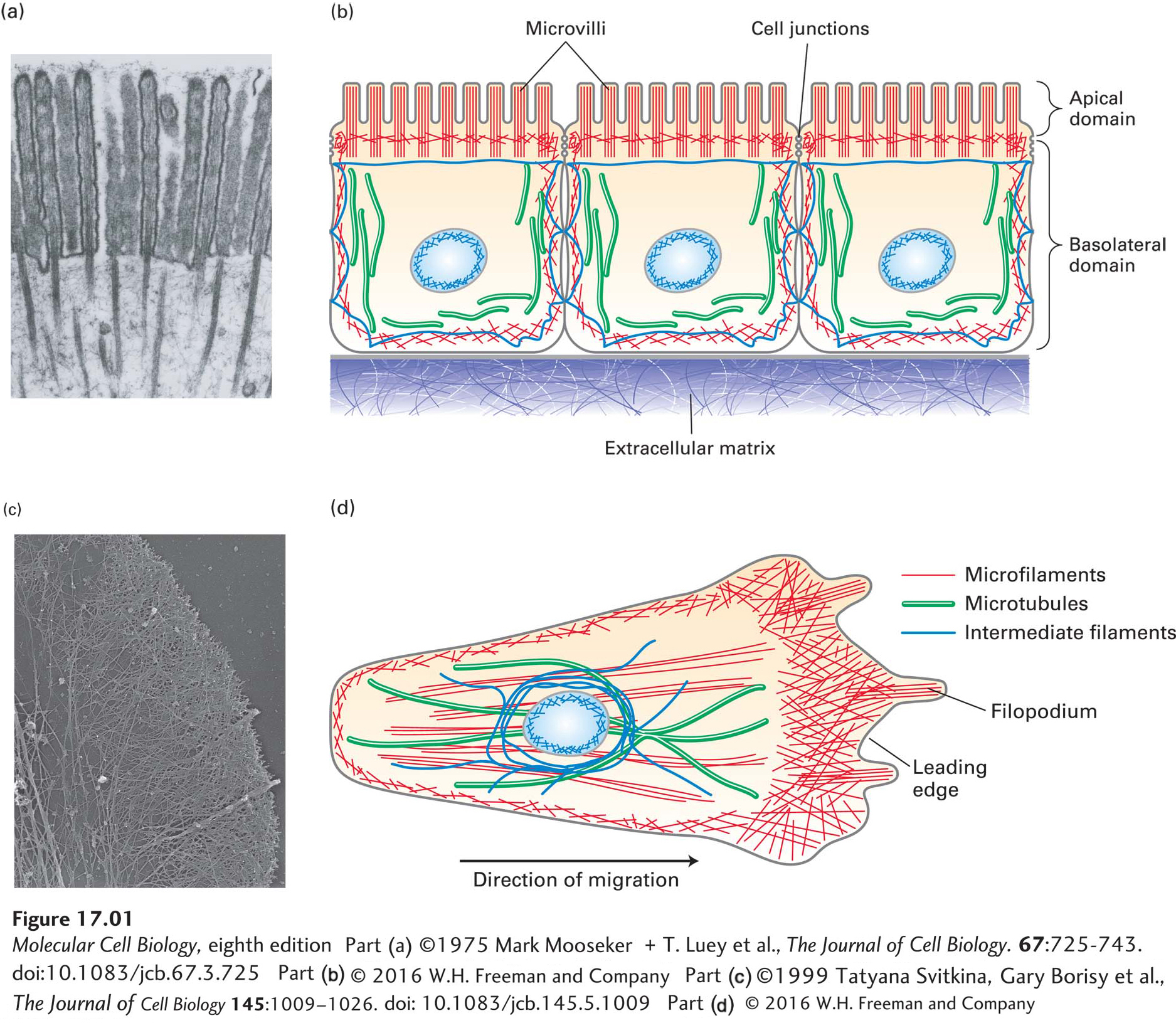
The epithelial cells that line the intestine form a tight, pavement-like layer of brick-shaped cells, known as an epithelium (Figure 17-1a, b). Their function is to import nutrients (such as glucose) from the intestinal lumen across the apical (top) plasma membrane and export them across the basolateral (bottom-side) plasma membrane toward the bloodstream. To perform this directional transport, the apical and basolateral plasma membranes of epithelial cells must have different protein compositions. Epithelial cells are attached and sealed together by cell junctions (discussed in Chapter 20), which create a physical barrier between the apical and basolateral domains of the membrane. This separation allows the cell to place the correct transport proteins in the plasma membranes of the two surfaces. In addition, the apical membrane has a unique morphology, with numerous fingerlike projections called microvilli that increase the area of the plasma membrane available for nutrient absorption. To achieve this organization, epithelial cells must have an internal structure to give them shape and to deliver the appropriate proteins to the correct membrane surface.
FIGURE 17-1Overview of the cytoskeletons of an epithelial cell and a migrating cell. (a) Transmission electron micrograph of a thin section of an epithelial cell from the small intestine, showing the core bundles of microfilaments that provide support to the microvilli. (b) Epithelial cells are highly polarized, with distinct apical and basolateral domains. An intestinal epithelial cell transports nutrients into the cell through the apical domain and out of the cell across the basolateral domain. (c) Transmission electron micrograph of part of the leading edge of a migrating cell. The cell was treated with a mild detergent to dissolve the membranes, which also allows solubilization of most cytoplasmic components. The remaining cytoskeleton was shadowed with platinum and visualized in the electron microscope. Note the network of actin filaments visible in this micrograph. (d) A migrating cell, such as a fibroblast or a macrophage, has morphologically distinct domains, with a leading edge at the front. Microfilaments are indicated in red, microtubules in green, and intermediate filaments in dark blue. The position of the nucleus (light blue oval) is also shown.
Now consider the macrophage, a type of white blood cell that seeks out infectious agents and destroys them by an engulfing process called phagocytosis. Bacteria release chemicals that attract the macrophage and guide it to the infection. As the macrophage follows the chemical gradient, twisting and turning to get to the bacteria and phagocytose them, it must constantly reorganize its cell locomotion machinery. As we will see, the internal motile machinery of macrophages and other crawling cells is always oriented in the direction in which they crawl (Figure 17-1c, d).
These are just two examples of cell polarity: the ability of cells to generate functionally distinct regions. In fact, as you think about all types of cells, you will realize that most of them have some form of cell polarity. An additional and fundamental example of cell polarity is the ability of cells to divide: they must first select an axis for cell division and then set up the machinery to segregate their organelles along that axis.
Page 777
A cell’s shape, internal organization, and functional polarity are provided by a three-dimensional filamentous protein network called the cytoskeleton. The cytoskeleton can be isolated and visualized after treating cells with gentle detergents that solubilize the plasma membrane and internal organelles, releasing most of the cytoplasm (see Figure 17-1c). The cytoskeleton extends throughout the cell and is attached to the plasma membrane and internal organelles, thus providing a framework for cellular organization. The term cytoskeleton may imply a fixed structure like a bone skeleton. In fact, the cytoskeleton can be very dynamic, with components capable of reorganization in less than a minute, or it can be quite stable for hours at a time. As a result, the lengths and dynamics of cytoskeletal filaments can vary greatly, they can be assembled into diverse types of structures, and they can be regulated locally in the cell.
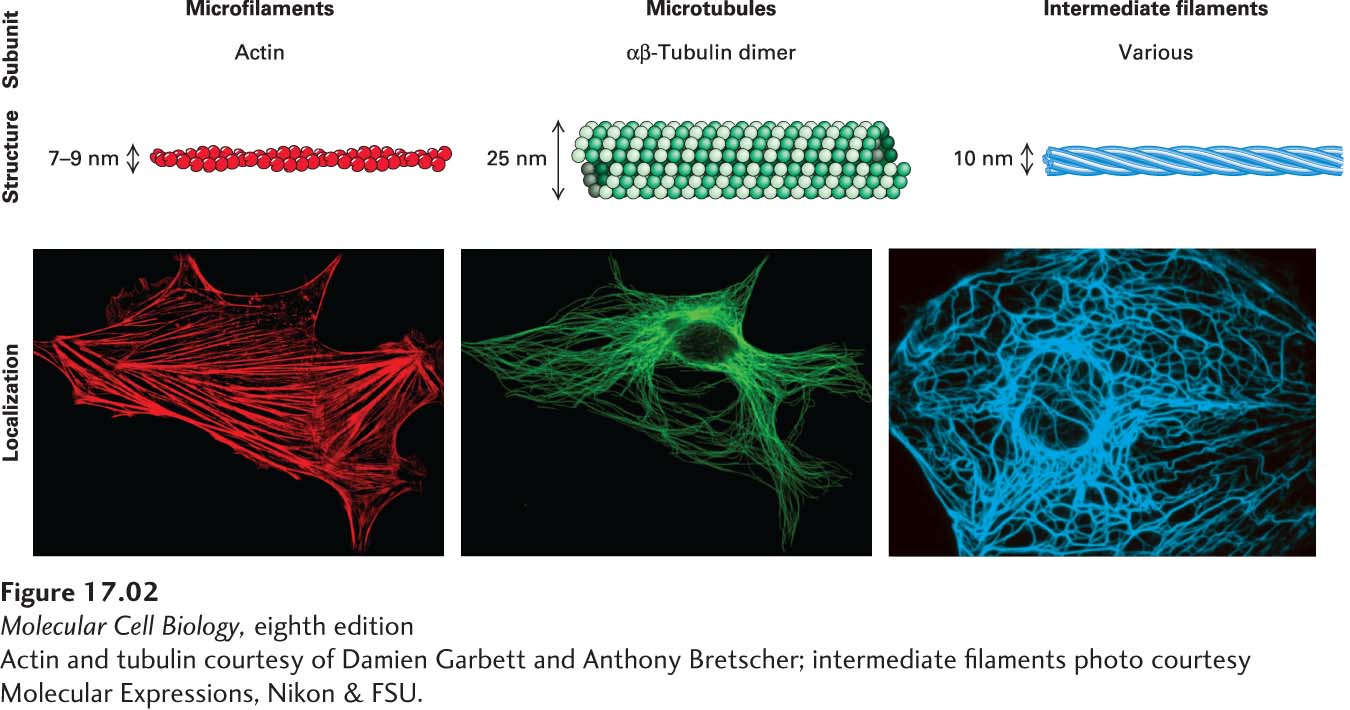
The cytoskeleton is composed of three major filament systems, shown in Figures 17-1b and 17-1d as well as in Figure 17-2. Each filament system is a polymer of assembled subunits that is organized and regulated in time and space. The subunits that make up the filaments undergo regulated assembly and disassembly, giving the cell the flexibility to assemble or disassemble different types of structures as needed.
[Actin and tubulin courtesy of Damien Garbett and Anthony Bretscher; intermediate filaments photo courtesy Molecular Expressions, Nikon & FSU.]
FIGURE 17-2The components of the cytoskeleton. Each filament type is assembled from specific subunits in a reversible process so that cells can assemble and disassemble filaments as needed. Bottom panels show localization of the three filament systems in cultured cells as seen by immunofluorescence microscopy of actin, tubulin, and an intermediate filament protein, respectively.
[Actin and tubulin courtesy of Damien Garbett and Anthony Bretscher; intermediate filaments photo courtesy Molecular Expressions, Nikon & FSU.]
Microfilaments are polymers of the protein actin that are organized into functional bundles and networks by actin-binding proteins. Microfilaments are especially important in the organization of the plasma membrane, giving shape to surface structures such as microvilli. Microfilaments can function on their own or serve as tracks for ATP-powered myosin motor proteins, which provide a contractile function (as in muscle) or ferry cargo along microfilaments.
Microtubules are long tubes formed by the protein tubulin and organized by microtubule-associated proteins. They often extend throughout the cell, providing an organizational framework for associated organelles and structural support to cilia and flagella. They also make up the structure of the mitotic spindle, the machine for separating duplicated chromosomes at mitosis. Motor proteins called kinesins and dyneins transport cargo along microtubules; like myosins, they are powered by ATP hydrolysis.
Intermediate filaments are tissue-specific filamentous structures that serve a number of different functions, including lending structural support to the nuclear membrane, providing structural integrity to cells in tissues, and serving structural and barrier functions in skin, hair, and nails. Unlike microfilaments and microtubules, intermediate filaments are not used as tracks by motor proteins.
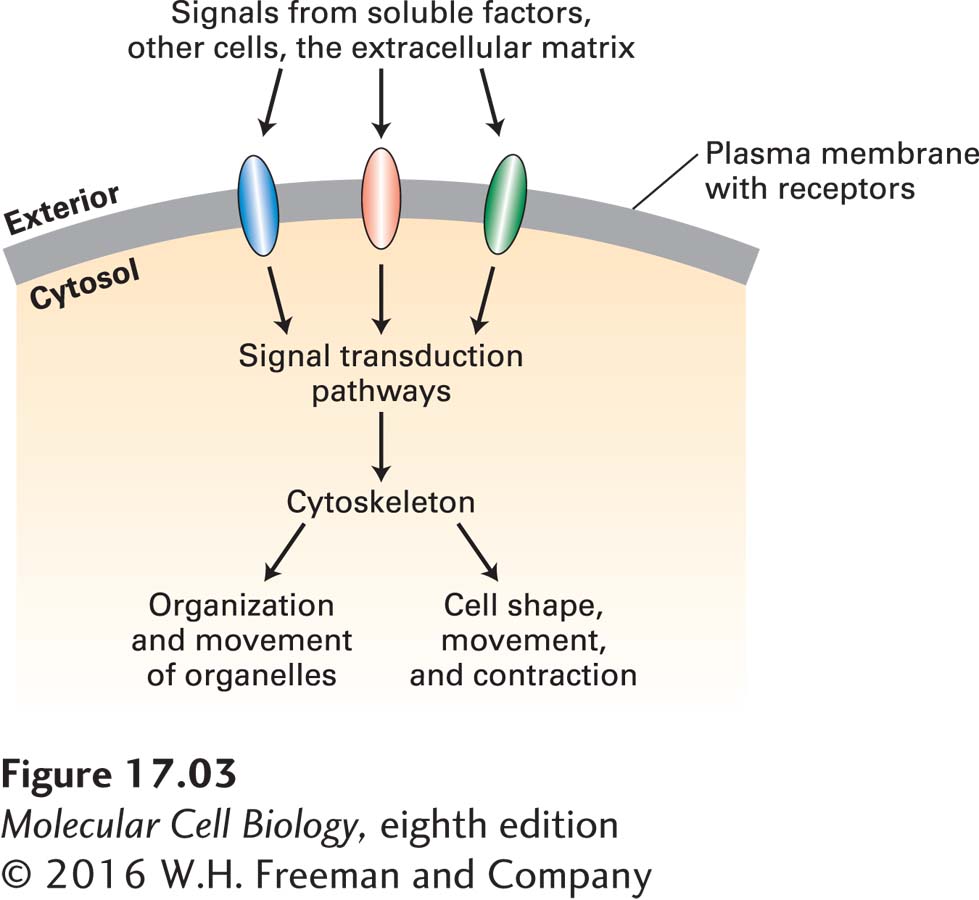
As we can see in Figure 17-1, different cells can construct very different arrangements of their cytoskeletons. To establish these arrangements, cells must sense signals—from soluble factors bathing the cell, from adjacent cells, or from the extracellular matrix—and interpret them (Figure 17-3). These signals are detected by cell-surface receptors that activate signal transduction pathways that ultimately converge on factors that regulate cytoskeletal organization.
FIGURE 17-3Regulation of cytoskeleton function by cell signaling. Cells use cell-surface receptors to sense external signals from the extracellular matrix, other cells, or soluble factors. These signals are transmitted across the plasma membrane and activate specific cytosolic signaling pathways. Signals—often integrated from more than one receptor—lead to the organization of the cytoskeleton so as to provide cells with their shape as well as to determine organelle distribution and movement. In the absence of external signals, cells still organize their internal structure, but not in a polarized manner.
The importance of the cytoskeleton for normal cell function and motility is evident when a defect in a cytoskeletal component—or in cytoskeletal regulation—causes a disease. For example, about 1 in 500 people has a defect that affects the contractile apparatus of the heart, which results in cardiomyopathies varying in degree of severity. Many diseases of red blood cells affect the cytoskeletal components that support these cells’ plasma membranes. Metastatic cancer cells exhibit unregulated motility due to misregulation of the cytoskeleton, breaking away from their tissue of origin and migrating to new locations to form new colonies of uncontrolled growth.
Page 778
In this and the following chapter, we discuss the structure, function, and regulation of the cytoskeleton. We see how a cell arranges its cytoskeleton to determine cell shape and polarity, to provide organization and motility to its organelles, and to be the structural framework for such processes as cell swimming and cell crawling. We discuss how cells assemble the three different filament systems and how signal transduction pathways regulate these structures both locally and globally. How the cytoskeleton is regulated during the cell cycle is discussed in Chapter 19, and how it participates in the functional organization of tissue is covered in Chapter 20. Our focus in this chapter is on microfilaments and actin-based structures. Although we initially examine the cytoskeletal systems separately, in the next chapter we will see that microfilaments cooperate with microtubules and intermediate filaments in the normal functioning of cells.