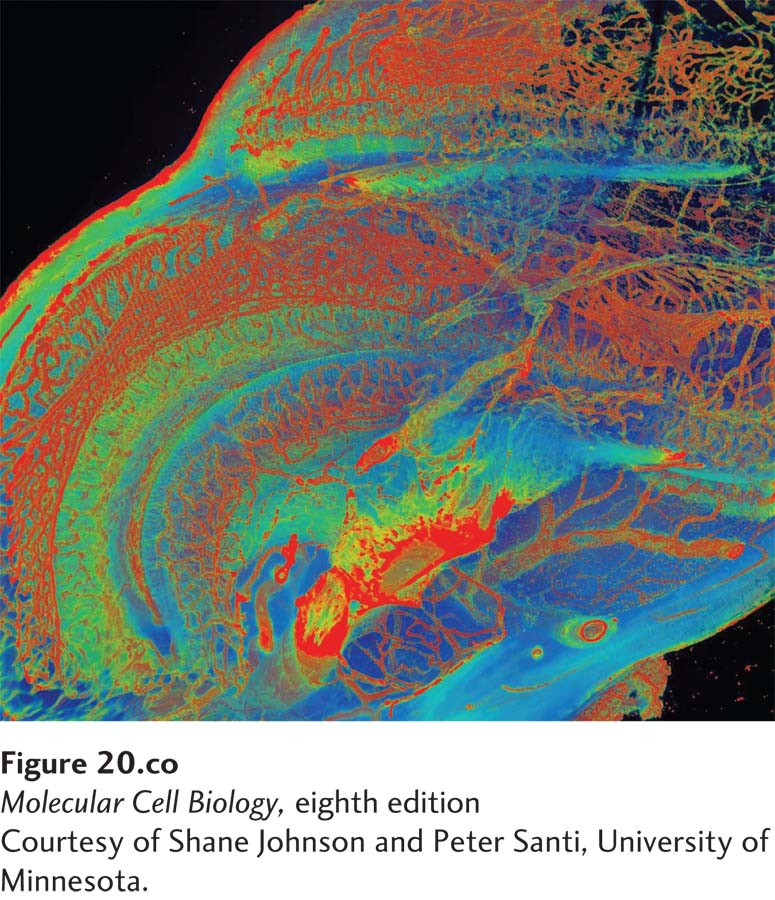
[Courtesy of Shane Johnson and Peter Santi, University of Minnesota.]
The cochlea of the inner ear uses mechanotransduction to convert the energy in sound waves into neuronal signals. The distribution of type IV collagen in the extracellular matrix of the cochlear duct of a mouse was visualized by scanning thin-sheet laser imaging microscopy after removing the cells with the detergent SDS and the calcium with the chelator EDTA. The sample was then stained first with an anti–type IV collagen antibody and then a fluorescently labeled secondary antibody. The false colors in the image represent the relative intensities of fluorescence (red > yellow > blue), and thus the relative local amounts of type IV collagen, in the basal lamina of the blood vessels (red), other basement membranes (yellow), and the cochlear wall (blue).
[Courtesy of Shane Johnson and Peter Santi, University of Minnesota.]
In the development of complex multicellular organisms such as plants and animals, progenitor cells differentiate into distinct “types” that have characteristic compositions, structures, and functions. Cells of a given type often aggregate into a tissue to cooperatively perform a common function: muscle contracts; neural tissue conducts electric impulses; xylem tissue in plants transports water. Different tissues can be organized into an organ, again to perform one or more specific functions. For instance, the muscles, valves, and blood vessels of a heart work together to pump blood. The coordinated functioning of many types of cells and tissues permits the organism to move, metabolize, reproduce, and carry out other essential activities. Indeed, the complex and diverse morphologies of plants and animals are examples of the whole being greater than the sum of the individual parts, more technically described as the emergent properties of a complex system.
Vertebrates have hundreds of different cell types, including leukocytes (white blood cells) and erythrocytes (red blood cells), photoreceptors in the retina, fat-storing adipocytes, fibroblasts in connective tissue, and the hundreds of different subtypes of neurons in the human brain. Even simple animals exhibit complex tissue organization. The adult form of the roundworm Caenorhabditis elegans contains a mere 959 cells, yet these cells fall into 12 different general cell types and many distinct subtypes. But despite their diverse forms and functions, all animal cells can be classified as components of just five main classes of tissue: epithelial tissue, connective tissue, muscular tissue, neural tissue, and blood. Various cell types are arranged in precise patterns of staggering complexity to generate tissues and organs. The costs of such complexity include increased requirements for information, material, energy, and time during the development of an individual organism. Although the physiological costs of complex tissues and organs are high, they confer the ability to thrive in varied and variable environments—a major evolutionary advantage.
Page 922
One of the defining characteristics of animals such as ourselves with complex tissues and organs (metazoans) is that the external and internal surfaces of most of their tissues and organs—and indeed, the exterior of the entire organism—are built from tightly packed sheet-like layers of cells known as epithelia. The formation of an epithelium and its subsequent remodeling into more complex collections of epithelial and nonepithelial tissues is a hallmark of the development of metazoans. Sheets of tightly attached epithelial cells act as regulatable, selectively permeable barriers, which permit the generation of chemically and functionally distinct compartments in an organism, such as the stomach and bloodstream. As a result, distinct and sometimes opposite functions (e.g., digestion and synthesis) can efficiently proceed simultaneously within an organism. Such compartmentalization also permits more sophisticated regulation of diverse biological functions. In many ways, the roles of complex tissues and organs in an organism are analogous to those of organelles and membranes in individual cells.
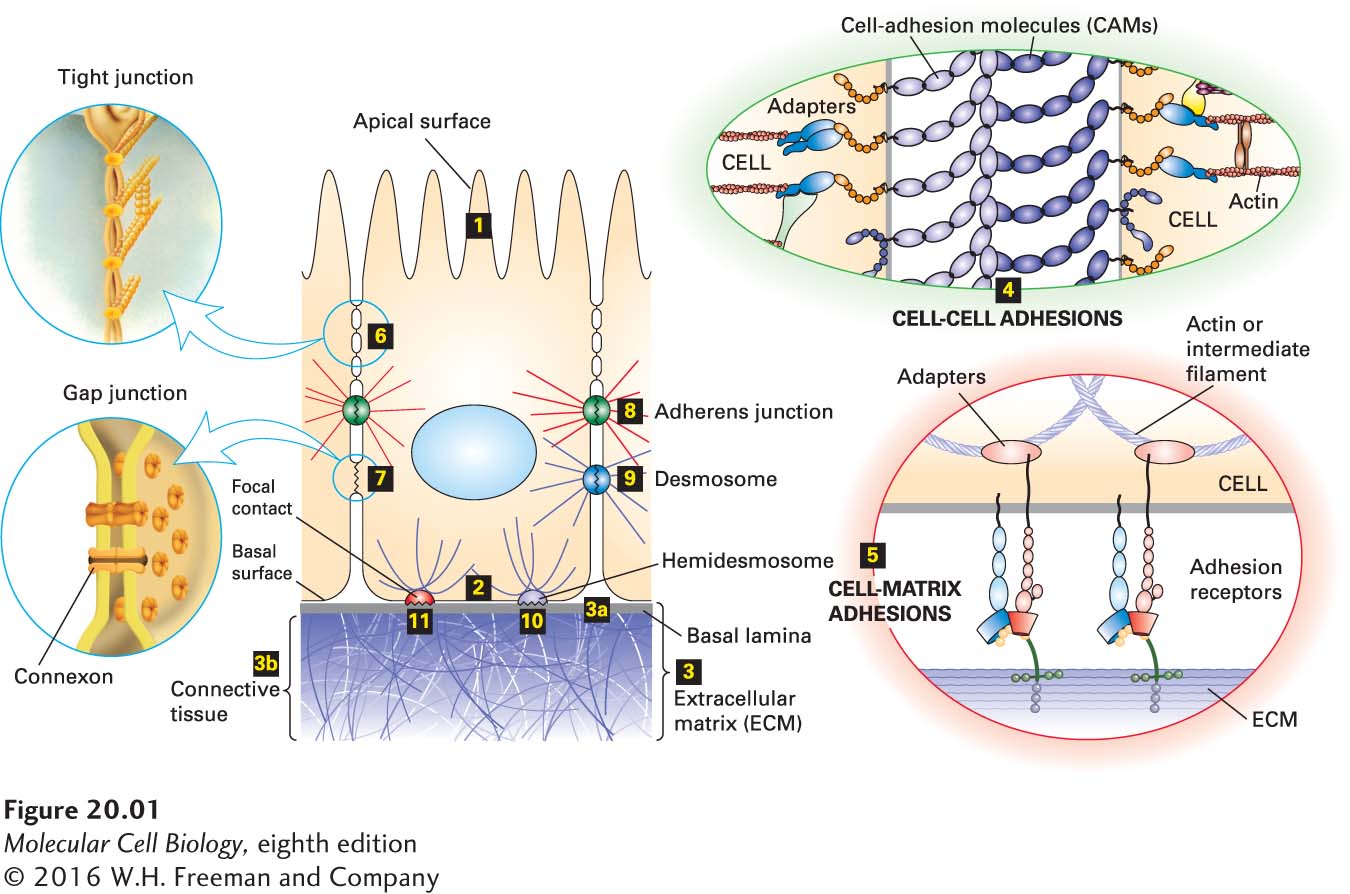
The assembly of distinct tissues and their organization into organs are determined by molecular interactions at the cellular level (Figure 20-1). These interactions would not be possible without the temporally, spatially, and functionally regulated expression of a wide array of adhesion molecules. Cells in tissues can adhere directly to one another (cell-cell adhesion) through specialized membrane proteins called cell-adhesion molecules (CAMs), which often cluster into specialized cell junctions. In the fruit fly Drosophila melanogaster, at least 500 genes (~4 percent of the total) are estimated to be involved in cell adhesion, and in mammals there are over 1000 such genes. Cells in animal tissues also adhere indirectly (cell-matrix adhesion) through the binding of adhesion receptors in the plasma membrane to components of the surrounding extracellular matrix (ECM), a complex interdigitating meshwork of proteins and polysaccharides secreted by cells into the spaces between them. Some adhesion receptors can also function as CAMs, mediating direct interaction between cells.
FIGURE 20-1Overview of major cell-cell and cell-matrix adhesive interactions. Schematic cutaway drawing of a typical epithelial tissue, such as in the inner surface of the intestines. The apical (upper) surface of each cell is packed with fingerlike microvilli (1) that project into the intestinal lumen, and the basal (lower) surface (2) rests on extracellular matrix (ECM). The ECM (3) associated with epithelial cells is usually organized into various interconnected layers—such as the basal lamina (3a), connecting fibers (not shown), and connective tissue (3b)—in which large, interdigitating ECM macromolecules bind to one another and to the cells (3). Cell-adhesion molecules (CAMs) bind to CAMs on other cells, mediating cell-cell adhesion (4), and adhesion receptors bind to various components of the ECM, mediating cell-matrix adhesion (5). Both types of cell-surface adhesion molecules are usually integral membrane proteins whose cytosolic domains often bind to multiple intracellular adapter proteins. These adapters, directly or indirectly, link the CAM to the cytoskeleton (actin or intermediate filaments) and to intracellular signaling pathways (as illustrated in Figure 20-8). As a consequence, information can be transferred by CAMs and the macromolecules to which they bind from the cell exterior to the intracellular environment (outside-in) and vice versa (inside-out). In some cases, a complex aggregate of CAMs, adapters, and associated proteins is assembled. Specific localized aggregates of CAMs or adhesion receptors form various types of cell junctions, which play important roles in holding tissues together and facilitating communication between cells and their environment. Tight junctions (6), lying just under the apical surface, prevent the diffusion of many substances through the extracellular spaces between the cells. Through connexon channels, gap junctions (7) allow the movement of small molecules and ions between the cytosols of adjacent cells. The remaining three types of junctions, adherens junctions (8 and 4), desmosomes (9), hemidesmosomes (10 and 5), and focal contacts (also called focal adhesions; 11) link the cytoskeleton of a cell to other cells or to the ECM. See V. Vasioukhin and E. Fuchs, 2001, Curr. Opin. Cell Biol.13:76–84.
Page 923
Cell-cell and cell-matrix adhesions not only allow cells to aggregate into distinct tissues, but also provide a means for the bidirectional transfer of information between the exterior and the interior of cells. As we will see, both types of adhesions are intrinsically associated with the cytoskeleton and cellular signaling pathways. As a result, a cell’s surroundings influence its shape and functional properties (“outside-in” effects); likewise, cellular shape and function influence a cell’s surroundings (“inside-out” effects). Thus connectivity and communication are intimately related properties of cells in tissues. Information transfer is important to many biological processes, including cell survival, proliferation, differentiation, and migration. Therefore, it is not surprising that defects that interfere with adhesive interactions and the associated flow of information can cause or contribute to diseases, including a wide variety of neuromuscular and skeletal disorders and cancer.
In this chapter, we examine various types of adhesion molecules found on the surfaces of cells and in the surrounding extracellular matrix. Interactions between these molecules allow the organization of cells into tissues and have profound effects on tissue development, function, and pathology. Many adhesion molecules are members of families or superfamilies of related proteins. While each type of adhesion molecule performs a distinct role, we will focus on the common features shared by members of some of these families to illustrate the general principles underlying their structures and functions. Because of the particularly well-understood nature of the adhesion molecules in tissues that form tight epithelia, as well as their very early evolutionary development, we will initially focus on epithelial tissues, such as the walls of the intestinal tract and the skin. Epithelial cells are normally nonmotile (sessile); however, during development, wound healing, and in certain pathological states (e.g., cancer), epithelial cells can transform into motile cells. Changes in the expression and function of adhesion molecules play a key role in this transformation, as they do in normal biological processes involving cell movement, such as the crawling of white blood cells into sites of infection. We therefore follow the discussion of epithelial tissues with a discussion of adhesion in nonepithelial, developing, and motile tissues.
The evolutionary lineages of plants and animals diverged before multicellular organisms arose. Thus multicellularity and the molecular means for assembling tissues and organs must have arisen independently in animal and plant lineages. Not surprisingly, then, animals and plants exhibit many differences in the organization and development of tissues. For this reason, we first consider the organization of tissues in animals and then deal separately with plants.