We turn now to the assembly of plant cells into tissues. The overall structural organization of plants is generally simpler than that of animals. For instance, plants have only four broad types of cells, which in mature plants form four basic classes of tissue: dermal tissue interacts with the environment, vascular tissue transports water and dissolved substances such as sugars and ions, space-filling ground tissue constitutes the major sites of metabolism, and sporogenous tissue forms the reproductive organs. Plant tissues are organized into just four main organ systems: stems have support and transport functions, roots provide anchorage and absorb and store nutrients, leaves are the sites of photosynthesis, and flowers enclose the reproductive structures. Thus, at the cell, tissue, and organ levels, plants are generally less complex than most animals.
Moreover, unlike animals, plants do not replace or repair old or damaged cells or tissues; they simply grow new organs. Indeed, the developmental fate of any given plant cell is primarily based on its position in the organism rather than on its lineage, whereas both are important in animals (see Chapter 21). In both plants and animals, a cell’s direct communication with its neighbors is important. Most important for this chapter, and in contrast with animals, few cells in plants contact one another directly through molecules incorporated into their plasma membranes. Instead, plant cells are typically surrounded by a rigid cell wall that contacts the cell walls of adjacent cells (Figure 20-41a). Also in contrast with animal cells, a plant cell rarely changes its position in the organism relative to other cells. These features of plants and their organization have determined the distinctive molecular mechanisms by which plant cells are incorporated into tissues and communicate with one another.
[Part (b) courtesy Maureen C. McCann. Part (c) republished with permission of Oxford University Press, from Fujino, T., et al., “Characterization of cross-links between cellulose microfibrils, and their occurrence during elongation growth in pea epicotyl,” Plant Cell Physiol. 2000, 41(4):486–94; permission conveyed through the Copyright Clearance Center, Inc.]
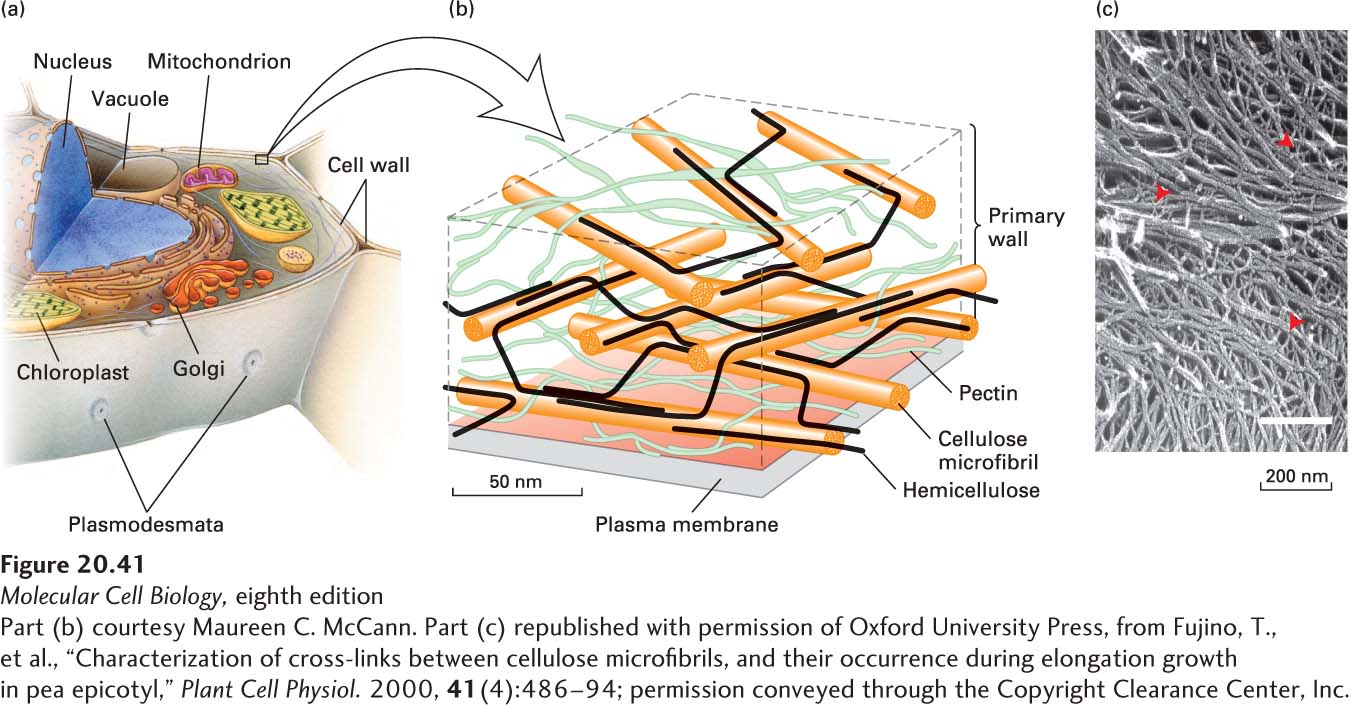
FIGURE 20-41Structure of the plant cell wall. (a) Overview of the organization of a typical plant cell, in which the organelle-filled cell with its plasma membrane is surrounded by a well-defined extracellular matrix called the cell wall. (b) Schematic representation of the cell wall of an onion. Cellulose and hemicellulose are arranged into at least three layers in a matrix of pectin. The sizes of the polymers and their separations are drawn to scale. To simplify the diagram, most of the hemicellulose cross-links and other matrix constituents (e.g., extensin, lignin) are not shown. See M. McCann and K. R. Roberts, 1991, in C. Lloyd, ed., The Cytoskeletal Basis of Plant Growth and Form, Academic Press, p. 126. (c) Quick-freeze deep-etch electron micrograph of the cell wall of a garden pea in which some of the pectin molecules were removed by chemical treatment. The abundant thicker fibers are cellulose microfibrils, and the thinner fibers are hemicellulose cross-links (red arrowheads).
[Part (b) courtesy Maureen C. McCann. Part (c) republished with permission of Oxford University Press, from Fujino, T., et al., “Characterization of cross-links between cellulose microfibrils, and their occurrence during elongation growth in pea epicotyl,” Plant Cell Physiol. 2000, 41(4):486–94; permission conveyed through the Copyright Clearance Center, Inc.]