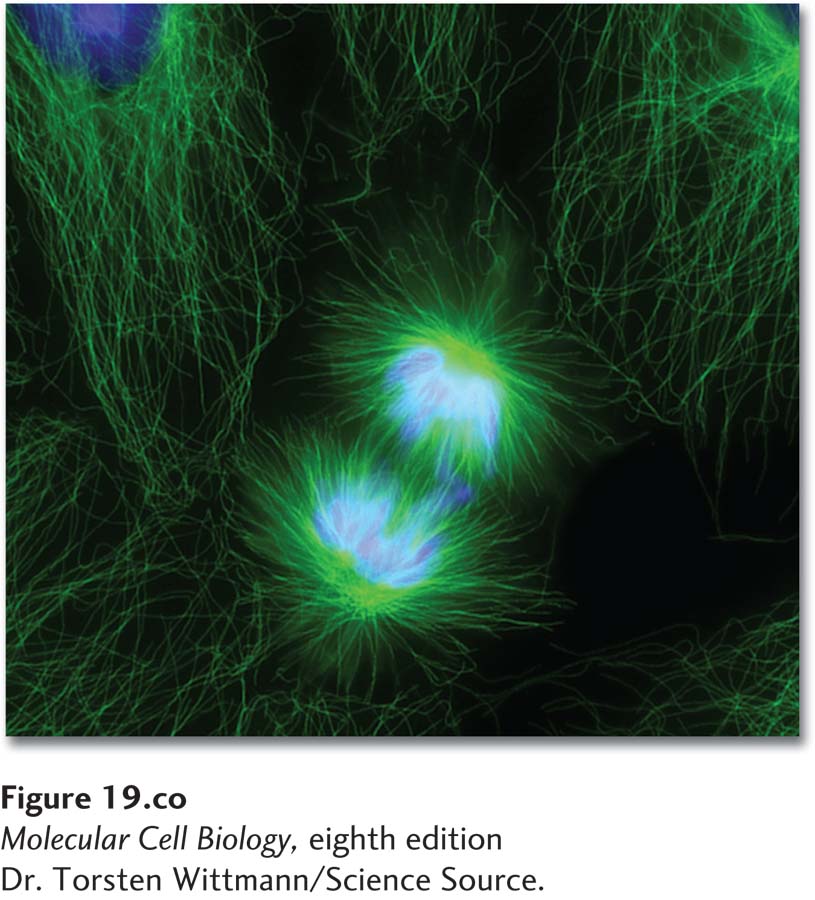
Micrograph of a human epithelial cell undergoing anaphase. Following DNA replication, cells undergo mitosis to segregate their replicated chromosomes. The cell in the center is in the process of segregating its chromosomes (blue) to opposite ends using the mitotic spindle apparatus (green). This process occurs during anaphase. Thereafter, the cytoplasm of the cell is divided to produce two identical daughter cells.
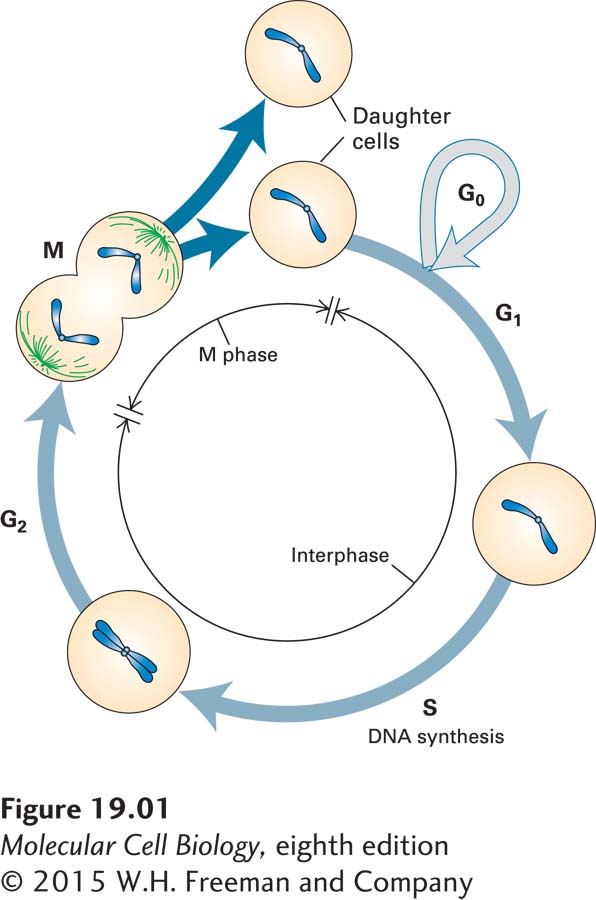
FIGURE 19-1The fate of a single parent chromosome throughout the eukaryotic cell cycle. Following mitosis (M), daughter cells contain 2n chromosomes in diploid organisms and 1n chromosomes in haploid organisms. In proliferating cells, G1 is the period between the “birth” of a cell following mitosis and the initiation of DNA synthesis, which marks the beginning of the S phase. At the end of the S phase, cells enter G2 containing twice the number of chromosomes they had as G1 cells (4n in diploid organisms, 2n in haploid organisms). The end of G2 is marked by the onset of mitosis, during which numerous events leading to cell division occur. The G1, S, and G2 phases are collectively referred to as interphase, the period between one mitosis and the next. Most nonproliferating cells in vertebrates leave the cell cycle in G1, entering the G0 state. Although chromosomes condense only during mitosis, here they are shown in condensed form throughout the cell cycle to emphasize the number of chromosomes at each stage. For simplicity, the nuclear envelope is not depicted.
Proper control of cell division is vital to all organisms. In unicellular organisms, cell division must be balanced with cell growth so that cell size is properly maintained. If several divisions occur before parent cells have reached the proper size, daughter cells eventually become too small to be viable. If cells grow too large before cell division, the cells function improperly and the number of cells increases slowly. In developing multicellular organisms, the replication of each cell must be precisely controlled and timed to faithfully and reproducibly complete the developmental program in every individual. Each type of cell in every tissue must control its replication precisely for normal development of complex organs such as the brain or the kidney. In a normal adult, cells divide only when and where they are needed. However, loss of normal controls on cell replication is the fundamental defect in cancer, an all-too-familiar disease that kills one in every six people in the developed world (see Chapter 24). The molecular mechanisms regulating eukaryotic cell division discussed in this chapter have gone a long way in explaining how replication control goes awry in cancer cells. Appropriately, Leland Hartwell, Tim Hunt, and Paul Nurse were awarded the Nobel Prize in Physiology or Medicine in 2001 for the initial experiments that elucidated the master regulators of cell division in all eukaryotes.
The term cell cycle refers to the ordered series of events that lead to cell division and the production of two daughter cells, each containing chromosomes identical to those of the parent cell. Two main molecular processes take place during the cell cycle, with resting intervals in between: during the S phase of the cycle, each parent chromosome is duplicated to form two identical sister chromatids; and in mitosis (M phase), the resulting sister chromatids are distributed to each daughter cell (Figure 19-1; see also Figure 1-16). Chromosome replication and segregation to daughter cells must occur in the proper order in every cell division. If a cell undergoes chromosome segregation before the replication of all chromosomes has been completed, at least one daughter cell will lose genetic information. Likewise, if a second round of replication occurs in one region of a chromosome before cell division occurs, the genes encoded in that region are increased in number out of proportion to other genes, a phenomenon that often leads to an imbalance of gene expression that is incompatible with viability.
Page 874
High accuracy and fidelity are required to ensure that DNA replication is carried out correctly and that each daughter cell inherits the correct number of each chromosome. To achieve this, cell division is controlled by surveillance mechanisms known as checkpoint pathways, which prevent initiation of each step in cell division until the earlier steps on which it depends have been completed and any mistakes that occurred during the process have been corrected. Mutations that inactivate or alter the normal operation of these checkpoint pathways contribute to the generation of cancer cells because they result in chromosomal rearrangements and abnormal numbers of chromosomes, which lead to further mutations and changes in gene expression that cause uncontrolled cell growth (see Chapter 24).
In the late 1980s, it became clear that the molecular processes regulating the two key events in the cell cycle—chromosome replication and chromosome segregation—are fundamentally similar in all eukaryotic cells. Initially, it was surprising to many researchers that cells as diverse as budding yeast and developing human neurons use nearly identical proteins to regulate their division. However, like transcription and protein synthesis, control of cell division appears to be a fundamental cellular process that evolved and was largely optimized early in eukaryotic evolution. Because of this similarity, research with diverse organisms, each with its own particular experimental advantages, has contributed to a growing understanding of how cell cycle events are coordinated and controlled. Biochemical, genetic, imaging, and micromanipulation techniques have all been employed in studying various aspects of the eukaryotic cell cycle. These studies have revealed that cell division is controlled primarily by regulation of the timing of entry into the cell division cycle, DNA replication, and mitosis.
The master controllers of the cell cycle are a small number of protein kinases, each of which contains a regulatory subunit (cyclin) and a catalytic subunit (cyclin-dependent kinase, or CDK). These heterodimeric kinases regulate the activities of multiple proteins involved in entry into the cell cycle, DNA replication, and mitosis by phosphorylating those proteins at specific regulatory sites, activating some and inhibiting others to coordinate their activities. Regulated degradation of proteins also plays a prominent role in important cell cycle transitions. Since protein degradation is irreversible, it ensures that the processes move in only one direction through the cell cycle.
In this chapter, we first present an overview of the cell cycle and then describe the various experimental systems that have contributed to our current understanding of it. We then discuss cyclin-dependent kinases (CDKs) and the many different ways in which these key cell cycle controllers are regulated. Next we examine each cell cycle phase in greater detail, with an emphasis on how control of CDK activity governs the events that take place in each phase. We then discuss the checkpoint pathways that establish the order of the cell cycle and ensure that each cell cycle phase occurs with accuracy. The chapter concludes with a discussion of meiosis, a special type of cell division that generates haploid germ cells (eggs and sperm), and the molecular mechanisms that distinguish it from mitosis. In our discussion, we emphasize the general principles governing cell cycle progression and use a species-spanning nomenclature when discussing the factors controlling each cell cycle phase.