Pre-tRNAs Undergo Extensive Modification in the Nucleus
Mature cytosolic tRNAs, which average 75–80 nucleotides in length, are produced from larger precursors (pre-tRNAs) synthesized by RNA polymerase III in the nucleoplasm. Mature tRNAs contain numerous modified bases that are not present in tRNA primary transcripts. Cleavage and base modification occur during processing of all pre-tRNAs; some pre-tRNAs are also spliced during processing. All of these processing and modification events occur in the nucleus.
Page 467
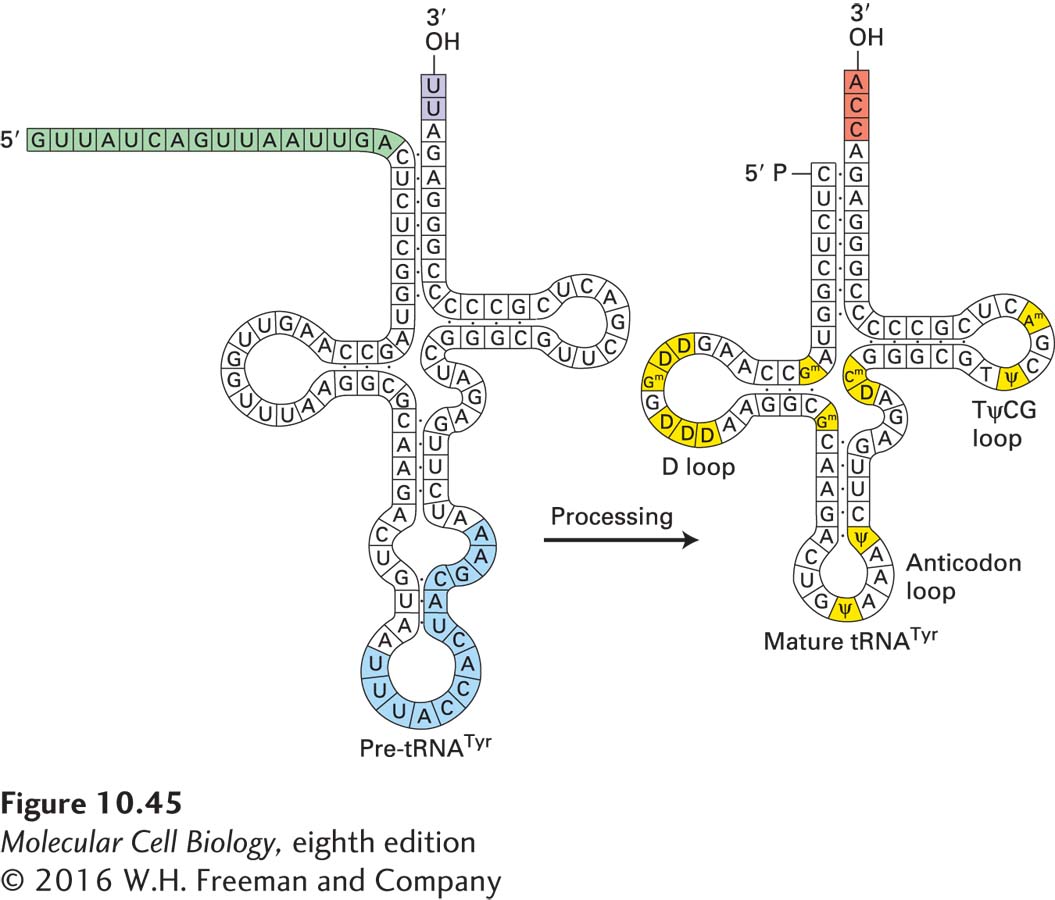
A 5′ sequence of variable length that is absent from mature tRNAs is present in all pre-tRNAs (Figure 10-45). These extra 5′ nucleotides are present because the 5′ end of a mature tRNA is generated by an endonucleolytic cleavage specified by the tRNA three-dimensional structure, rather than by the start site of transcription. The extra nucleotides are removed by ribonuclease P (RNase P), a ribonucleoprotein endonuclease. Studies with E. coli RNase P indicate that at high Mg2+ concentrations, its RNA component alone can recognize and cleave E. coli pre-tRNAs. The RNase P polypeptide increases the rate of cleavage by the RNA, allowing cleavage to proceed at physiological Mg2+ concentrations. A comparable RNase P functions in eukaryotes.
About 10 percent of the bases in pre-tRNAs are modified enzymatically during processing. Three classes of base modifications occur (see Figure 10-45):
U residues at the 3′ end of pre-tRNA are replaced with a CCA sequence. The CCA sequence is found at the 3′ end of all tRNAs and is required for their charging by aminoacyl-tRNA synthetases during protein synthesis. This step in tRNA synthesis probably functions as a quality-control point, since only properly folded tRNAs are recognized by the CCA addition enzyme.
Methyl and isopentenyl groups are added to the heterocyclic ring of purine bases, and the 2′-OH groups in the ribose of specific residues are methylated.
Specific uridines are converted to dihydrouridine, pseudouridine, or ribothymidine residues. The functions of these base and ribose modifications are not well understood, but since they are highly conserved, they probably have a positive influence on protein synthesis.
FIGURE 10-45Changes that occur during the processing of tyrosine pre-tRNA. A 14-nucleotide intron (blue) in the anticodon loop is removed by splicing. A 16-nucleotide sequence (green) at the 5′ end is cleaved by RNase P. U residues at the 3′ end are replaced by the CCA sequence (red) found in all mature tRNAs. Numerous bases in the stem-loops are converted to characteristic modified bases (yellow). Not all pre-tRNAs contain introns that are spliced out during processing, but they all undergo the other types of changes shown here. D = dihydrouridine; Ψ = pseudouridine.
As shown in Figure 10-45, the pre-tRNA expressed from the yeast tyrosine tRNA (tRNATyr) gene contains a 14-base intron that is not present in mature tRNATyr. Some other eukaryotic tRNA genes and some archaeal tRNA genes also contain introns. The introns in nuclear pre-tRNAs are shorter than those in pre-mRNAs and lack the consensus splice-site sequences found in pre-mRNAs (see Figure 10-7). Pre-tRNA introns are also clearly distinct from the much longer self-splicing group I and group II introns found in chloroplast and mitochondrial pre-rRNAs. The mechanism of pre-tRNA splicing differs in three fundamental ways from the mechanisms used by self-splicing introns and spliceosomes (see Figure 10-44). First, splicing of pre-tRNAs is catalyzed by proteins, not by RNAs. Second, a pre-tRNA intron is excised in one step that entails simultaneous cleavage at both ends of the intron. Finally, hydrolysis of GTP and ATP is required to join the two tRNA halves generated by cleavage on either side of the intron.
After pre-tRNAs are processed in the nucleoplasm, the mature tRNAs are transported to the cytoplasm through nuclear pore complexes by exportin-t, an exportin (see Chapter 13) dedicated to the nuclear export of tRNAs. In the cytoplasm, tRNAs are passed between aminoacyl-tRNA synthetases, elongation factors, and ribosomes during protein synthesis (see Chapter 5). Thus tRNAs are generally associated with proteins and spend little time free in the cell, as is also the case for mRNAs and rRNAs.