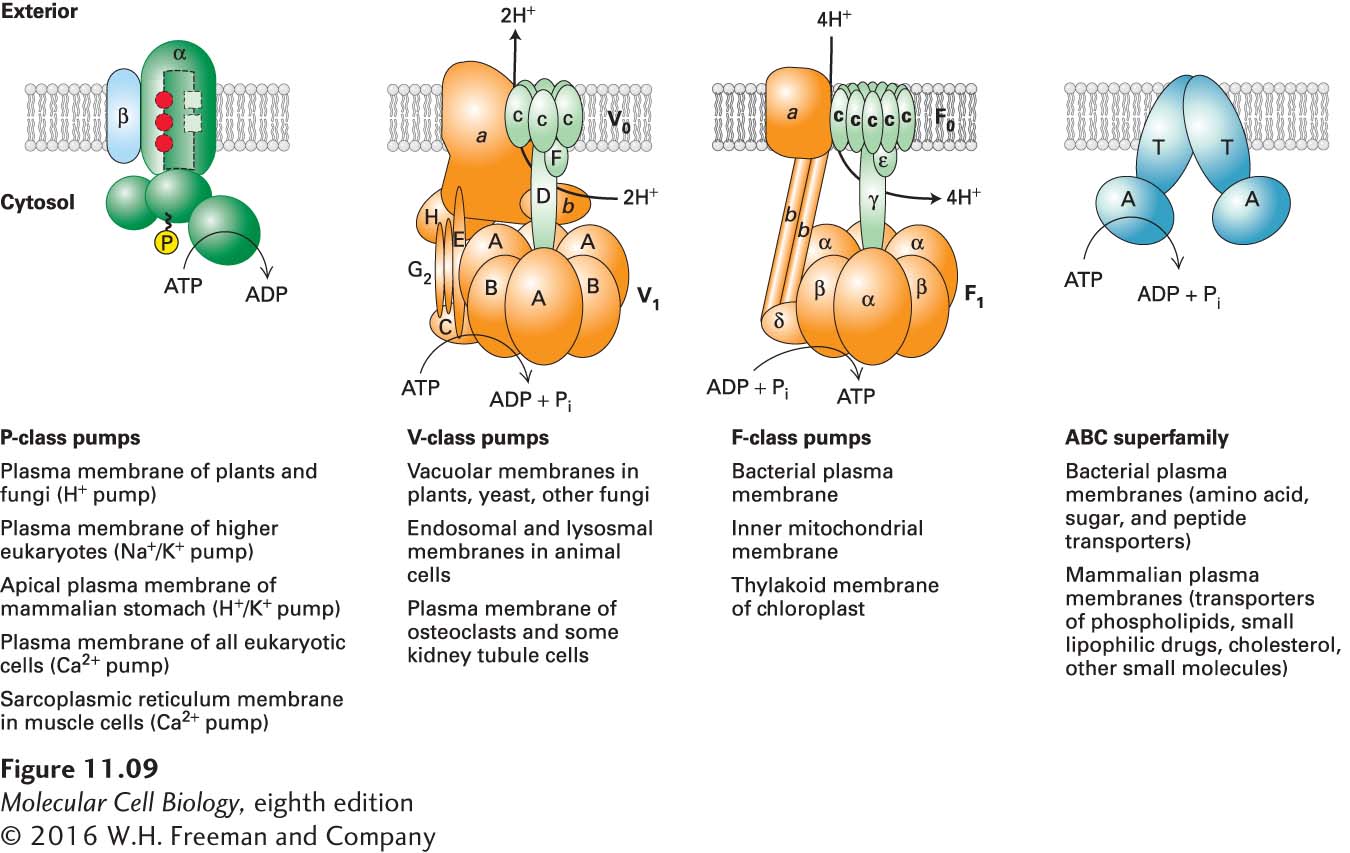
The general structures of the four classes of ATP-powered pumps are depicted in Figure 11-9, with specific examples in each class listed below the figure. Note that the members of three of the classes (P, F, and V) transport only ions, as do some members of the fourth class, the ABC superfamily. Most members of the ABC superfamily, however, transport small molecules such as amino acids, sugars, peptides, lipids, and many types of drugs.
FIGURE 11-9The four classes of ATP-powered transport proteins. The locations of specific examples are indicated below each class. P-class pumps are composed of two catalytic α subunits, which become phosphorylated as part of the transport cycle. Two β subunits, present in some of these pumps, may regulate transport. Only one α and one β subunit are depicted here. V-class and F-class pumps do not form phosphoprotein intermediates, and almost all transport only protons. Their structures are similar and contain similar proteins, but none of their subunits are related to those of P-class pumps. V-class pumps couple ATP hydrolysis to transport of protons against a concentration gradient, whereas F-class pumps normally operate in the reverse direction and use the energy in a proton concentration or voltage gradient to synthesize ATP. All members of the large ABC superfamily of proteins contain two transmembrane (T) domains and two cytosolic ATP-binding (A) domains, which couple ATP hydrolysis to solute movement. These core domains are present as separate subunits in some ABC proteins (as depicted here) but are fused into a single polypeptide in other ABC proteins. See T. Nishi and M. Forgac, 2002, Nat. Rev. Mol. Cell Biol.3:94; C. Toyoshima et al., 2000, Nature405:647; D. McIntosh, 2000, Nat. Struct. Biol.7:532; and T. Elston, H. Wang, and G. Oster, 1998, Nature391:510.
All P-class pumps possess two identical catalytic α subunits, each of which contains an ATP-binding site. Most also have two smaller β subunits, which usually have regulatory functions. During transport, at least one of the α subunits becomes phosphorylated (hence the name “P” class), and the transported ions move through the phosphorylated subunit. The amino acid sequences around the phosphorylated residues are homologous in different pumps. This class includes the Na+/K+ ATPase in the plasma membrane, which generates the low cytosolic Na+ and high cytosolic K+ concentrations typical of animal cells (see Figure 11-3). Certain Ca2+ ATPases pump Ca2+ ions out of the cytosol into the external medium; others pump Ca2+ from the cytosol into the endoplasmic reticulum or into the specialized ER called the sarcoplasmic reticulum that is found in muscle cells. Another member of the P class, found in acid-secreting cells of the mammalian stomach, transports protons (H+ ions) out of and K+ ions into the cell.
Page 485
The structures of V-class and F-class pumps are similar to one another but are unrelated to, and more complicated than, those of P-class pumps. V- and F-class pumps contain several different transmembrane and cytosolic subunits. Virtually all known V and F pumps transport only protons and do so in a process that does not involve a phosphoprotein intermediate. V-class pumps generally function to generate the low pH of plant vacuoles and of lysosomes and other acidic vesicles in animal cells by pumping protons from the cytosolic to the exoplasmic face of the membrane against a proton electrochemical gradient. In contrast, the proton pumps that generate and maintain the plasma-membrane electric potential in plant, fungal, and many bacterial cells belong to the P class of pumps.
F-class pumps are found in bacterial plasma membranes and in mitochondria and chloroplasts. In contrast to V-class pumps, they generally function as reverse proton pumps, in which the energy released by the energetically favored movement of protons from the exoplasmic to the cytosolic face of the membrane down the proton electrochemical gradient is used to power the energetically unfavorable synthesis of ATP from ADP and Pi. Because of their importance in ATP synthesis in chloroplasts and mitochondria, F-class proton pumps, commonly called ATP synthases, are treated separately in Chapter 12 (Cellular Energetics).
The final class of ATP-powered pumps is a large family with multiple members that are more diverse in function than those of the other classes. Referred to as the ABC (ATP-binding cassette) superfamily, this class includes several hundred different transport proteins found in organisms ranging from bacteria to humans. As detailed below, some of these transport proteins were first identified as multidrug-resistance proteins that, when overexpressed in cancer cells, export anticancer drugs and render tumors resistant to their action. Each ABC protein is specific for a single substrate or group of related substrates, which may be ions, sugars, amino acids, phospholipids, cholesterol, peptides, polysaccharides, or even proteins. All ABC membrane transport proteins share a structural organization consisting of four “core” domains: two transmembrane (T) domains, which form the passageway through which transported molecules cross the membrane, and two cytosolic ATP-binding (A) domains. In some ABC proteins, mostly those in bacteria, the core domains are present as four separate polypeptides; in others, the core domains are fused into one or two multidomain polypeptides. ATP binding and hydrolysis drives the transport process in most ABC membrane transporters. However, the energy from ATP binding and hydrolysis can also be used to regulate the opening and closing of a continuous channel, as described below for the cystic fibrosis transmembrane conductance regulator protein (CFTR).