[Dylan Burnette and Jennifer Lippincott-Schwartz.]
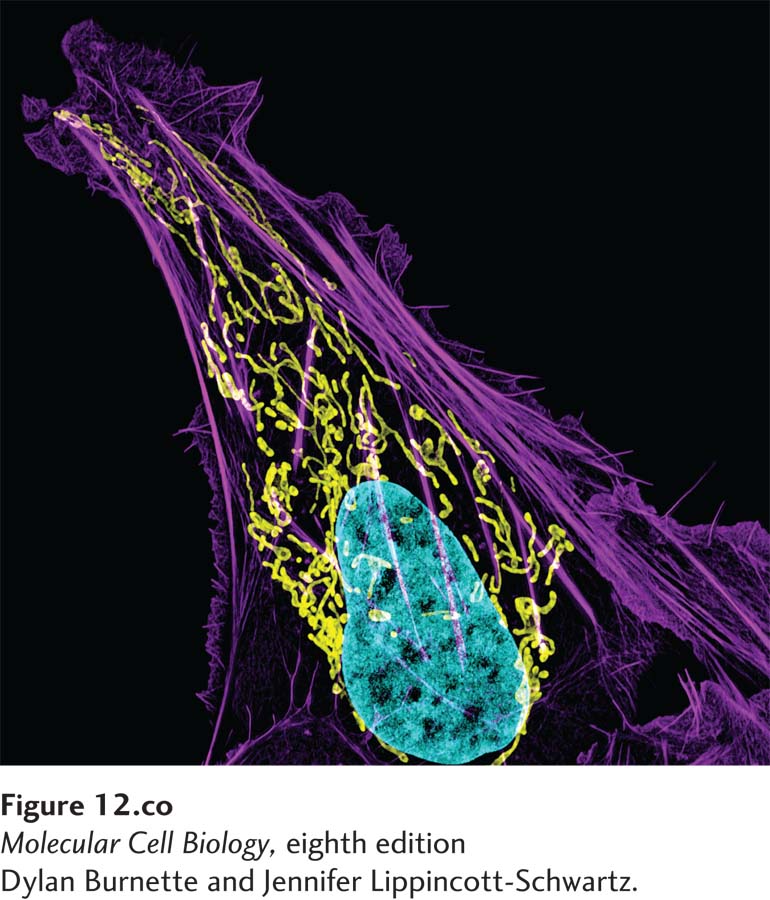
Fluorescence micrograph showing the yellow tubelike network of mitochondria in a human bone cancer (osteosarcoma) cell along with the DNA in the nucleus (aqua) and actin cytoskeletal fibers (purple).
[Dylan Burnette and Jennifer Lippincott-Schwartz.]
From the growth and division of a cell to the beating of a heart to the electrical activity of a neuron that underlies thinking, life requires energy. Energy is defined as the capacity to do work, and on a cellular level, that work includes conducting and regulating a multitude of chemical reactions and transport processes, growing and dividing, generating and maintaining a highly organized structure, and interacting with other cells. This chapter describes the molecular mechanisms by which cells use sunlight or chemical nutrients as sources of energy, with a special focus on how cells convert these external sources of energy into a biologically universal intracellular chemical energy carrier, adenosine triphosphate, or ATP (Figure 12-1). ATP, found in all types of organisms and presumably present in the earliest life forms, is generated by the chemical addition of inorganic phosphate (HPO42−, often abbreviated as Pi) to adenosine diphosphate, or ADP, a process called phosphorylation. Cells use the energy released during hydrolysis of the terminal phosphoanhydride bond in ATP (see Figure 2-31) to power many otherwise energetically unfavorable processes. Examples include the synthesis of proteins from amino acids and of nucleic acids from nucleotides (see Chapter 4), the transport of molecules against a concentration gradient by ATP-powered pumps (see Chapter 11), the contraction of muscles (see Chapter 17), and the beating of cilia (see Chapter 18). A key theme of cellular energetics is that proteins use, or “couple,” energy released from one process (e.g., ATP hydrolysis) to drive another process (e.g., movement of molecules across membranes) that otherwise would be thermodynamically unfavorable.
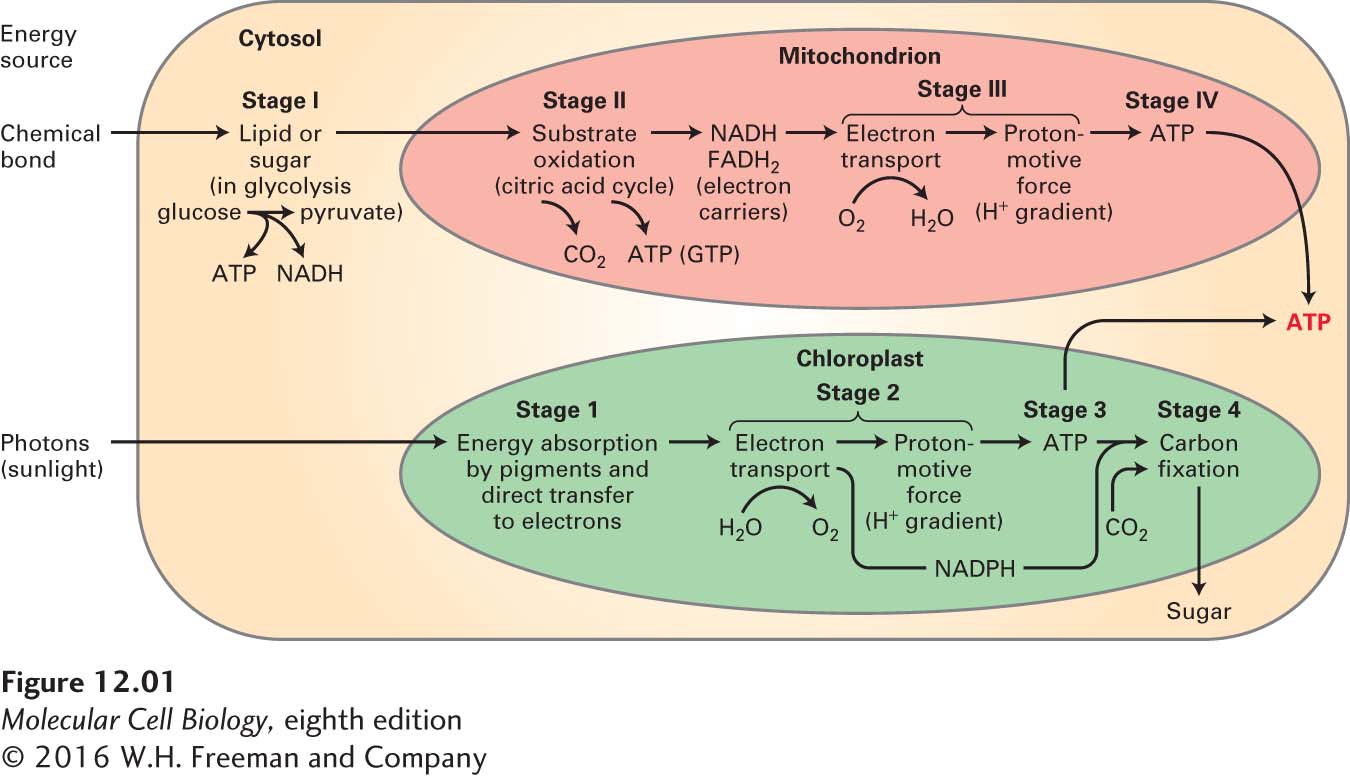
FIGURE 12-1Overview of aerobic oxidation and photosynthesis. Eukaryotic cells use two fundamental mechanisms to convert external sources of energy into ATP. (Top) In aerobic oxidation, “fuel” molecules [primarily sugars and fatty acids (lipids)] undergo preliminary processing in the cytosol, such as breakdown of glucose to pyruvate (stage I), and are then transferred into mitochondria, where they are converted by oxidation with O2 to CO2 and H2O (stages II and III) and ATP is generated (stage IV). (Bottom) In photosynthesis, which occurs in chloroplasts, the radiant energy of light is absorbed by specialized pigments (stage 1); the absorbed energy is used both to oxidize H2O to O2 and to establish conditions (stage 2) necessary for the generation of ATP (stage 3) and of carbohydrates from CO2 (carbon fixation, stage 4). Both mechanisms involve the production of reduced high-energy electron carriers (NADH, NADPH, FADH2) and the movement of electrons down an electric potential gradient in an electron-transport chain through specialized membranes. Energy released from these electrons is captured as a proton electrochemical gradient (proton-motive force) that is then used to drive ATP synthesis. Bacteria use comparable processes.
The energy to drive ATP synthesis from ADP (ΔG°′ = 7.3 kcal/mol) derives primarily from two sources: the energy in the chemical bonds of nutrients and the energy in sunlight (see Figure 12-1). The two processes primarily responsible for converting these energy sources into ATP are aerobic oxidation (also known as aerobic respiration), which occurs in mitochondria in nearly all eukaryotic cells (see Figure 12-1, top), and photosynthesis, which occurs only in chloroplasts, found in the leaf cells of plants (see Figure 12-1, bottom) and in certain single-celled organisms, such as algae and cyanobacteria. Two additional processes, glycolysis and the citric acid cycle (see Figure 12-1, top), are also important direct or indirect sources of ATP in both animal and plant cells.
In aerobic oxidation, breakdown products of sugars (carbohydrates) and fatty acids (hydrocarbons)—both derived from the digestion of food in animals—are converted by oxidation with oxygen (O2) to carbon dioxide (CO2) and water (H2O). The energy released from this overall reaction is transformed into the chemical energy of phosphoanhydride bonds in ATP. This process is analogous to burning wood (carbohydrates) or oil (hydrocarbons) to generate heat in furnaces or motion in automobile engines: both consume O2 and generate CO2 and H2O. The key difference is that cells break the overall reaction down into many intermediate steps, with the amount of energy released in any given step closely matched to the amount of energy that can be stored—for example, as ATP—or that is required for the next intermediate step. If there were not such a close match, excess released energy would be lost as heat (which would be very inefficient), or not enough energy would be released to generate energy storage molecules such as ATP or to drive the next step in the process (which would be ineffective).
Page 514
In photosynthesis, the radiant energy of light is absorbed by pigments such as chlorophyll and used to make ATP and carbohydrates—primarily sucrose and starch. Unlike aerobic oxidation, which uses carbohydrates and O2 to generate CO2, photosynthesis uses CO2 as a substrate and generates O2 and carbohydrates as products.
This reciprocal relationship between aerobic oxidation in mitochondria and photosynthesis in chloroplasts underlies a profound symbiotic relationship between photosynthetic and non-photosynthetic organisms. The oxygen generated during photosynthesis is the source of virtually all the oxygen in the air, and the carbohydrates produced are the ultimate source of energy for virtually all non-photosynthetic organisms on earth. (An exception is bacteria living in deep-sea hydrothermal vents—and the organisms that feed on them—which obtain energy for converting CO2 into carbohydrates by oxidation of geologically generated reduced inorganic compounds released by the vents.)
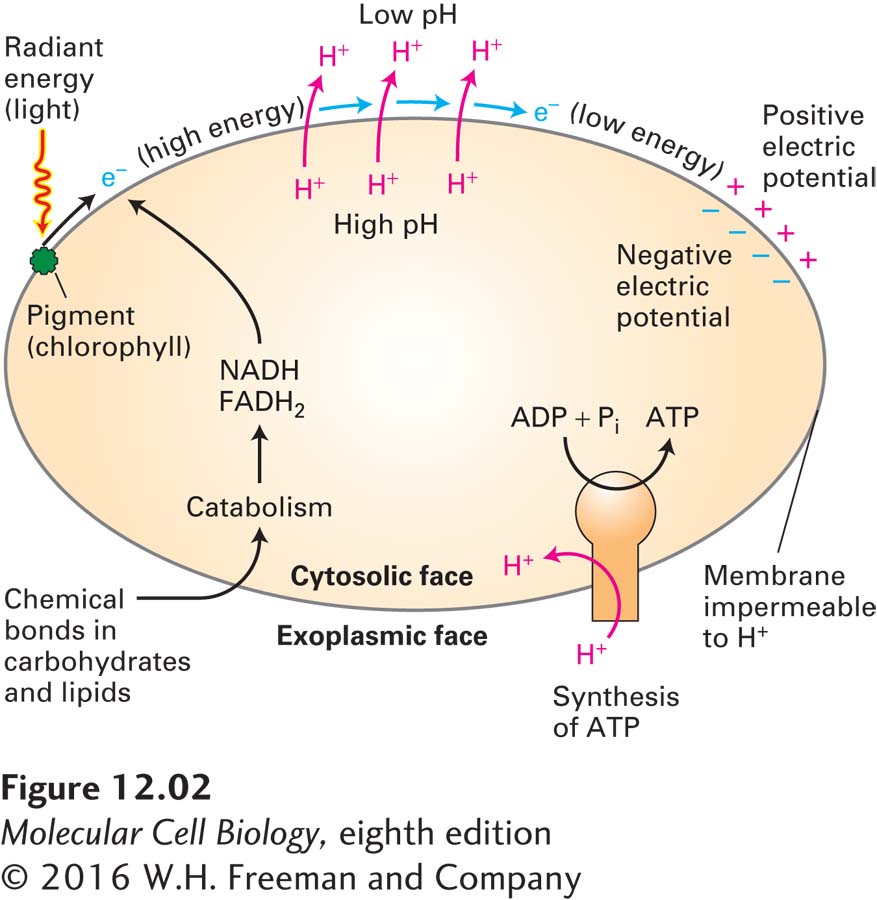
At first glance, it might seem that the molecular mechanisms of photosynthesis and aerobic oxidation have little in common, besides the fact that they both produce ATP. However, a revolutionary discovery in cell biology established that bacteria, mitochondria, and chloroplasts all use the same mechanism, known as chemiosmosis, to generate ATP from ADP and Pi. In chemiosmosis (also known as chemiosmotic coupling), a proton electrochemical gradient across a membrane is first generated by energy released as electrons travel down their electric potential gradient through an electron-transport chain. The energy stored in this proton electrochemical gradient, called the proton-motive force, is then used to power the synthesis of ATP (Figure 12-2) or other energy-requiring processes. As protons move down their electrochemical gradient through the ATP synthesizing enzyme called ATP synthase, ATP is generated from ADP and Pi, a process that is the reverse of that mediated by the ATP-powered ion pumps discussed in Chapter 11. In this chapter, we explore the molecular mechanisms of the two processes that share this central mechanism, focusing first on aerobic oxidation and then on photosynthesis.
Page 515
FIGURE 12-2The proton-motive force powers ATP synthesis. Transmembrane proton concentration and electrical (voltage) gradients, collectively called the proton-motive force, are generated during aerobic oxidation and photosynthesis in both eukaryotes and prokaryotes (bacteria). High-energy electrons generated by light absorption by pigments (e.g., chlorophyll), or held in the reduced form of electron carriers (e.g., NADH, FADH2) made during the catabolism of sugars and lipids, pass down an electron-transport chain (blue arrows), releasing energy throughout the process. The released energy is used to pump protons across the membrane (red arrows), generating the proton-motive force. In chemiosmotic coupling, the energy released when protons flow down the gradient through ATP synthase drives the synthesis of ATP. The proton-motive force can also power other processes, such as the transport of metabolites across the membrane against their concentration gradient and rotation of bacterial flagella.