ABC Proteins Export a Wide Variety of Drugs and Toxins from the Cell
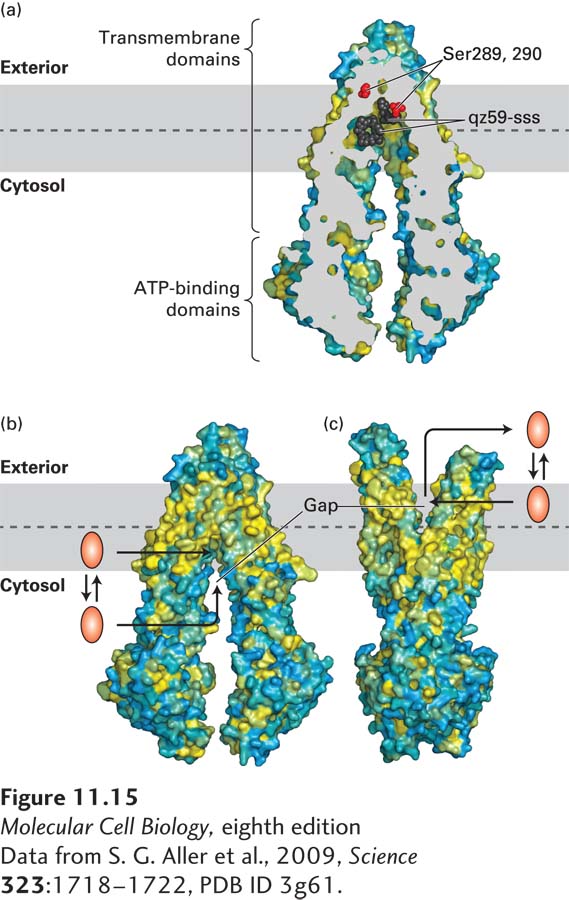
As noted earlier, all members of the very large and diverse ABC superfamily of membrane transport proteins contain two transmembrane (T) domains and two cytosolic ATP-binding (A) domains (see Figure 11-9). The T domains, each built of 10 membrane-spanning α helices, form the pathway through which the transported substance (substrate) crosses the membrane (Figure 11-15a) and determine the substrate specificity of each ABC protein. The sequences of the A domains are approximately 30–40 percent homologous in all members of this superfamily, indicating a common evolutionary origin.
[Data from S. G. Aller et al., 2009, Science323:1718–1722, PDB ID 3g61.]
FIGURE 11-15The multidrug transporter ABCB1 (MDR1): Structure and model of ligand export. (a) Cross-sectional view through the center of an ABCB1 protein bound to two molecules of a drug analog, qz59-sss (black), reveals the central location of the ligand-binding site in relation to the phospholipid bilayer: the central ligand-binding cavity is close to the leaflet-leaflet interface of the membrane. During transport, this binding cavity is alternately exposed to the exoplasmic and the cytosolic surface of the membrane. Serines 289 and 290 affect the ligand specificity of the transporter; they are shown as red spheres to highlight their juxtaposition to the bound ligand. Surface residues are colored yellow to denote hydrophobic and blue to denote hydrophilic amino acids. (b) Three-dimensional structure of ABCB1 with its ligand-binding site facing inward toward the cytosol. In this conformation, a hydrophilic ligand can bind directly from the cytosol. A more hydrophobic ligand can partition into the inner leaflet of the plasma membrane bilayer and then enter the ligand-binding site through a gap in the protein that is accessible directly from the hydrophobic core of the inner leaflet. (c) Model for the structure of ABCB1 with its ligand-binding site facing outward, based on the structures of homologous bacterial ABC proteins. When the protein assumes this conformation, the ligand can either diffuse into the exoplasmic leaflet or directly into the aqueous extracellular medium. See D. Gutman et al., 2009, Trends Biochem. Sci.35:36–42.
[Data from S. G. Aller et al., 2009, Science323:1718–1722, PDB ID 3g61.]
The first eukaryotic ABC protein to be recognized was discovered during studies on tumor cells and cultured cells that exhibited resistance to several drugs with unrelated chemical structures. Such cells were eventually shown to express elevated levels of a multidrug-resistance (MDR) transport protein originally called MDR1 and now known as ABCB1. This protein uses the energy derived from ATP hydrolysis to export a large variety of drugs from the cytosol to the extracellular medium. The Mdr1 gene is frequently amplified in multidrug-resistant cells, resulting in a large overproduction of the MDR1 protein. In contrast to bacterial ABC proteins, which are built of four discrete subunits, all four domains of mammalian ABCB1 are fused into a single 170-kDa protein.
The substrates of mammalian ABCB1 are primarily planar, lipid-soluble molecules with one or more positive charges; they all compete with one another for transport, which suggests that they bind to the same or overlapping sites on the protein. Many drugs transported by ABCB1 diffuse from the extracellular medium across the plasma membrane, unaided by transport proteins, into the cell cytosol, where they block various cellular functions. Two such drugs are colchicine and vinblastine, which block assembly of microtubules (see Chapter 18). ATP-powered export of such drugs by MDR1 reduces their concentration in the cytosol. As a result, a much higher extracellular drug concentration is required to kill cells that express ABCB1 than those that do not. That ABCB1 is an ATP-powered small-molecule pump has been demonstrated with liposomes containing the purified protein. Different drugs enhance the ATPase activity of these liposomes in a dose-dependent manner corresponding to their ability to be transported by ABCB1.
The three-dimensional structure of ABCB1, together with those of homologous bacterial ABC proteins, revealed the protein’s mechanism of transport as well as its ability to bind and transport a wide array of hydrophilic and hydrophobic substrates (see Figure 11-15). The two T domains form a binding site in the center of the membrane that alternates between an inward-facing (Figure 11-15b) and an outward-facing (Figure 11-15c) orientation, conforming to the alternating access model. The alternation between these two conformational states is powered by ATP binding to the two A subunits and its subsequent hydrolysis to ADP and Pi, but precisely how this happens is not known.
The substrate-binding cavity formed by ABCB1 is large. Some of the amino acids that line the cavity—mainly tyrosine and phenylalanine—have aromatic side chains, allowing ABCB1 to bind multiple types of hydrophobic ligands. Other segments of the cavity are lined with hydrophilic residues, allowing hydrophilic or amphipathic molecules to bind. In the inward-facing conformation, the binding site is open directly to the surrounding aqueous solution, allowing hydrophilic molecules to enter the binding site directly from the cytosol. In addition, a gap in the protein is accessible directly from the hydrophobic core of the inner leaflet of the membrane bilayer; this allows hydrophobic molecules to enter the binding site directly from the inner leaflet (see Figure 11-15b). After the ATP-powered change to the outward-facing conformation, molecules can exit the binding site into the outer membrane leaflet or directly into the extracellular medium (see Figure 11-15c).
Page 492
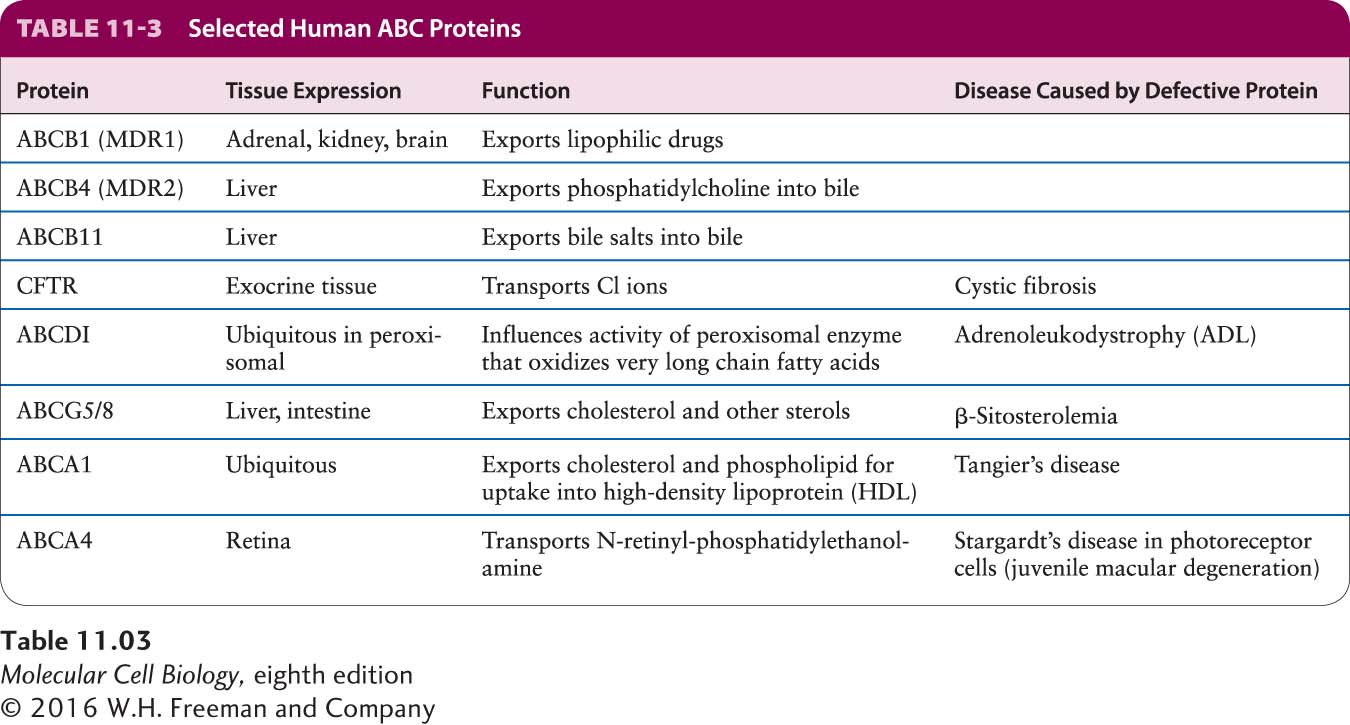
About 50 different mammalian ABC proteins are now recognized (Table 11-3). In eukaryotic cells, ABC proteins localize not only to the plasma membrane, but also to the membranes of many intracellular organelles. Several are expressed in abundance in the liver, intestines, and kidneys—sites where natural toxic and waste products are removed from the body. Substrates for these ABC proteins include sugars, amino acids, cholesterol, bile acids, phospholipids, peptides, proteins, toxins, and foreign substances. The normal function of ABCB1 most likely is to transport various natural and metabolic toxins into the bile or intestinal lumen for excretion or into the urine being formed in the kidney. During the course of its evolution, ABCB1 appears to have acquired the ability to transport drugs whose structures are similar to those of these endogenous toxins. Tumors derived from MDR-expressing cell types, such as hepatomas (liver cancers), are frequently resistant to virtually all chemotherapeutic agents and are thus difficult to treat, presumably because the tumors exhibit increased expression of ABCB1 or a related ABC protein.