Ion Channels Are Selective for Certain Ions by Virtue of a Molecular “Selectivity Filter”
All ion channels exhibit specificity for particular ions: K+ channels allow K+ ions, but not closely related Na+ ions, to enter them, whereas Na+ channels admit Na+, but not K+. Determination of the three-dimensional structure of a bacterial K+ channel first revealed how this exquisite ion selectivity is achieved. Comparisons of the sequences and structures of other K+ channels from organisms as diverse as bacteria, fungi, and humans established that all share a common structure and probably evolved from a single type of channel protein.
Page 498
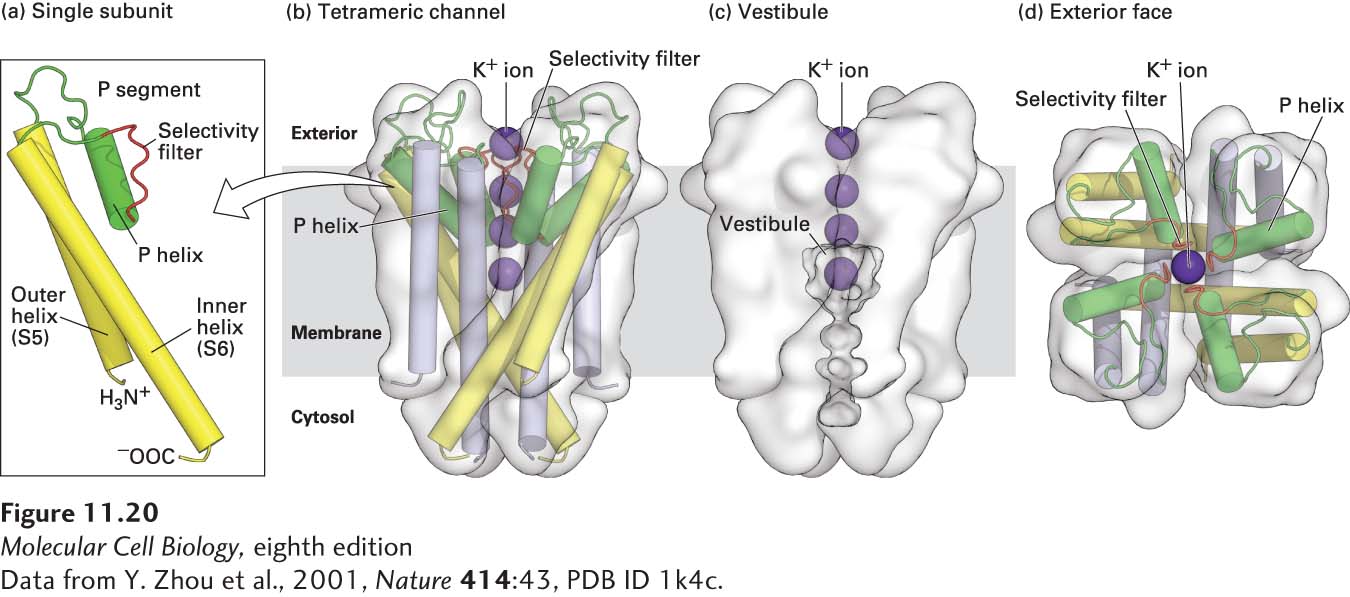
Like all other K+ channels, bacterial K+ channels are built of four identical transmembrane subunits symmetrically arranged around a central pore (Figure 11-20). Each subunit contains two membrane-spanning α helices (S5 and S6) and a short P (pore) segment that partly penetrates the membrane bilayer from the exoplasmic surface. In the tetrameric K+ channel, the eight transmembrane α helices (two from each subunit) form an inverted cone, generating a water-filled cavity called the vestibule in the central portion of the channel that extends halfway through the membrane toward the cytosolic side. Four extended loops that are part of the four P segments form the actual selectivity filter in the narrow part of the pore near the exoplasmic surface, above the vestibule.
[Data from Y. Zhou et al., 2001, Nature414:43, PDB ID 1k4c.]
FIGURE 11-20Structure of a resting K+ channel from the bacterium Streptomyces lividans. All K+ channels are tetramers comprising four identical subunits, each containing two conserved membrane-spanning α helices, called by convention S5 and S6, and a shorter P, or pore, segment. (a) One of the subunits, viewed from the side, with key structural features indicated. (b–d) The complete tetrameric channel viewed from the side (b and c) and from the top, or extracellular, end (d). The P segments (green) are located near the exoplasmic surface and connect the S5 and S6 α helices [yellow in (a), yellow and lavender in (b-d)]; they consist of a nonhelical “turret,” which lines the upper part of the pore; a short α helix; and an extended loop that protrudes into the narrowest part of the pore and forms the selectivity filter. This filter allows K+ (purple spheres), but not other ions, to pass. Below the filter is the central cavity, or vestibule, lined by the inner, or S6, α helices. The subunits in gated K+ channels, which open and close in response to specific stimuli, contain additional transmembrane helices not shown here; these channels are discussed in Chapter 22.
[Data from Y. Zhou et al., 2001, Nature414:43, PDB ID 1k4c.]
Several related pieces of evidence support the role of P segments in ion selection. First, the amino acid sequences of the P segments in all known K+ channels are highly homologous and are different from those in other ion channels. Second, mutations of certain amino acids in this segment alter the ability of a K+ channel to distinguish Na+ from K+. Finally, replacing the P segment of a bacterial K+ channel with the homologous segment from a mammalian K+ channel yields a chimeric protein that exhibits normal selectivity for K+ over other ions. Thus all K+ channels are thought to use the same mechanism to distinguish K+ from other ions.
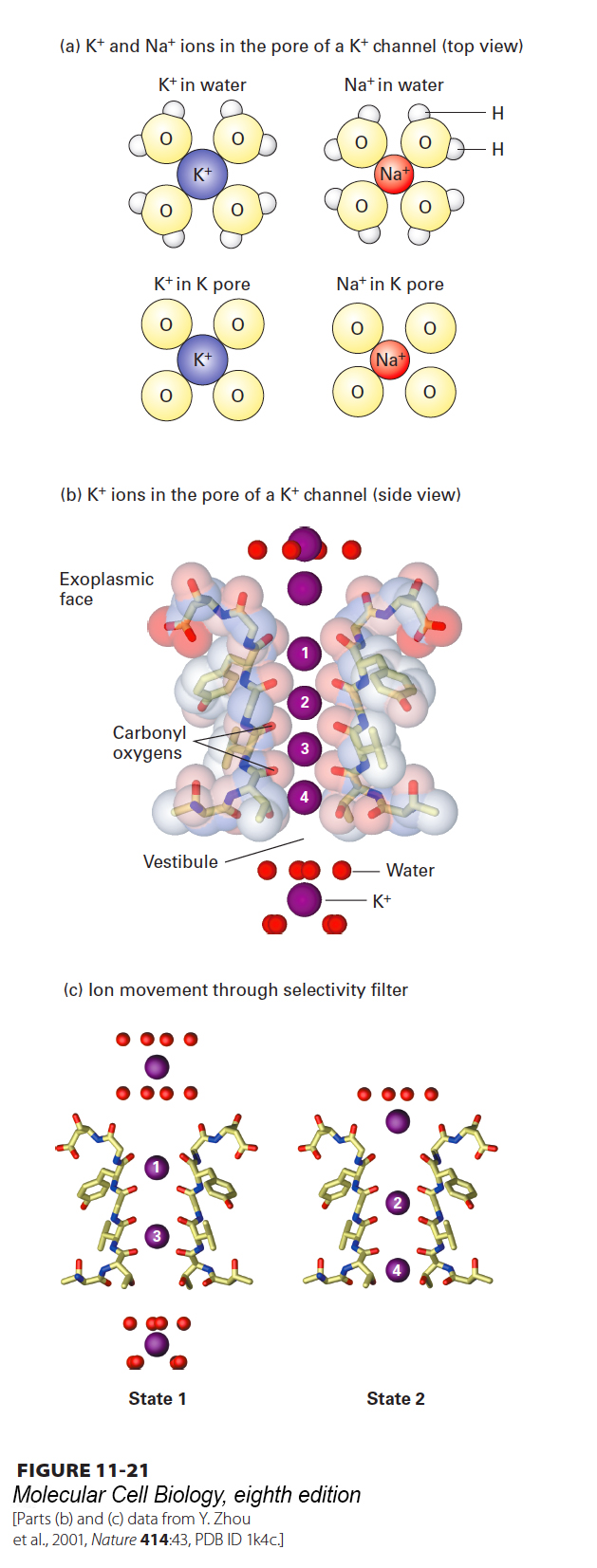
Na+ ions are smaller than K+ ions. How, then, can a channel protein exclude smaller Na+, yet allow passage of larger K+? The ability of the selectivity filter in K+ channels to select K+ over Na+ is due mainly to the backbone carbonyl oxygens on residues located in a Gly-Tyr-Gly sequence that is found in an analogous position in the P segment in every known K+ channel. As a K+ ion enters the selectivity filter—the narrow space between the P-segment filter sequences contributed by the four adjacent subunits—it loses its eight waters of hydration, but becomes bound in the same geometry to eight backbone carbonyl oxygens, two from the extended loop in each of the four P segments lining the channel (Figure 11-21a, bottom left). Thus little energy is required to strip off the eight waters of hydration of a K+ ion, and as a result, a relatively low activation energy is required for passage of K+ ions into the channel from an aqueous solution. A dehydrated Na+ ion is too small to bind to all eight carbonyl oxygens that line the selectivity filter with the same geometry as a Na+ ion surrounded by its normal eight water molecules in aqueous solution. As a result, Na+ ions “prefer” to remain in water rather than enter the selectivity filter, and thus the change in free energy for entry of Na+ ions into the channel is relatively high (Figure 11-21a, right). This difference in free energies favors passage of K+ ions through the channel over Na+ ions by a factor of 1000. Like Na+, the dehydrated Ca2+ ion is smaller than the dehydrated K+ ion and cannot interact properly with the oxygen atoms in the selectivity filter. Furthermore, because a Ca2+ ion has two positive charges and binds water oxygens more tightly than does a single-positive Na+ or K+ ion, more energy is required to strip the waters of hydration from Ca2+ than from K+ or Na+.
Page 499
[Parts (b) and (c) data from Y. Zhou et al., 2001, Nature414:43, PDB ID 1k4c.]
FIGURE 11-21Mechanism of ion selectivity and transport in resting K+ channels. (a) Schematic diagrams of K+ and Na+ ions hydrated in solution and in the pore of a K+ channel. As K+ ions pass through the selectivity filter, they lose their bound water molecules and become bound instead to eight backbone carbonyl oxygens (four of which are shown) that are part of the conserved amino acid sequence in the channel-lining selectivity filter loop of each P segment. The smaller Na+ ions, with their tighter shell of water molecules, cannot perfectly bind to the channel oxygen atoms and therefore pass through the channel only rarely. (b) High-resolution electron density map obtained from x-ray crystallography showing K+ ions (purple spheres) passing through the selectivity filter. Only two of the diagonally opposed channel subunits are shown. Within the selectivity filter, each unhydrated K+ ion interacts with eight carbonyl oxygen atoms (red sticks) lining the channel, two from each of the four subunits, as if to mimic the eight waters of hydration. (c) Interpretation of the electron density map, showing the two alternating states by which K+ ions move through the channel. Ion positions are numbered top to bottom from the exoplasmic side of the channel inward. In state 1, one sees a hydrated K+ ion with its eight bound water molecules, K+ ions at positions 1 and 3 within the selectivity filter, and a fully hydrated K+ ion within the vestibule. During K+ movement, each ion in state 1 moves one step inward, forming state 2. Thus in state 2, the K+ ion on the exoplasmic side of the channel has lost four of its eight waters, the ion at position 1 in state 1 has moved to position 2, and the ion at position 3 in state 1 has moved to position 4. In going from state 2 to state 1, the K+ at position 4 moves into the vestibule and picks up eight water molecules, while another hydrated K+ ion moves into the channel opening, and the other K+ ions move down one step. Note that K+ ions are shown here moving from the exoplasmic side of the channel to the cytosolic side because that is the normal direction of movement in bacteria. In animal cells, the direction of K+ movement is typically the reverse—from inside to outside. See C. Armstrong, 1998, Science280:56.
[Parts (b) and (c) data from Y. Zhou et al., 2001, Nature414:43, PDB ID 1k4c.]
Page 500
Recent x-ray crystallographic studies reveal that both when open and when closed, the K+ channel contains K+ ions within the selectivity filter; without these ions, the channel would probably collapse. The K+ ions are thought to be present either at positions 1 and 3 or at positions 2 and 4, each surrounded by eight carbonyl oxygen atoms (Figure 11-21b and c). Several K+ ions move simultaneously through the channel such that when the ion on the exoplasmic face that has been partially stripped of its water of hydration moves into position 1, the ion at position 2 jumps to position 3, and the one at position 4 exits the channel (Figure 11-21c).
The first three-dimensional structure of a sodium channel was determined in late 2011, and its mechanisms of ion selectivity are only now being determined. In contrast to potassium channels, all of which share a conserved selectivity filter, studies of sodium channels have revealed that distinct types of sodium channels in diverse species possess different types of selectivity filters, each of which uses a distinct mechanism to achieve selectivity for Na+ ions.