Three Main Classes of Membrane Proteins Transport Molecules and Ions Across Cellular Membranes
As is evident from Figure 11-1, very few molecules and no ions can cross a pure phospholipid bilayer at appreciable rates by simple diffusion. Thus transport of most molecules into and out of cells requires the assistance of specialized membrane proteins. Even in the cases of molecules with relatively large partition coefficients (e.g., urea, fatty acids) and certain gases, such as CO2 (carbon dioxide) and NH3 (ammonia), transport is frequently accelerated by specific proteins because simple diffusion does not occur rapidly enough to meet cellular needs.
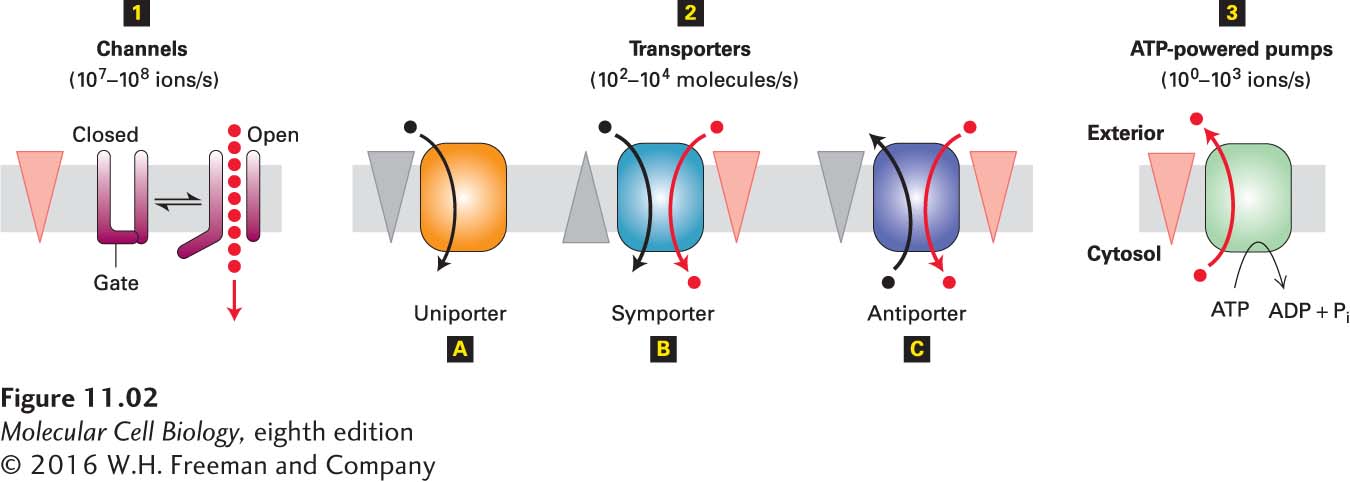
All membrane transport proteins are transmembrane proteins containing multiple membrane-spanning segments that are generally α helices. By forming a protein-lined pathway across the membrane, transport proteins are thought to allow hydrophilic substances to move through the membrane without coming into contact with its hydrophobic interior. Here we introduce the three main types of membrane transport proteins covered in this chapter (Figure 11-2).
FIGURE 11-2Overview of membrane transport proteins. Gradients are indicated by triangles with the tip pointing toward lower concentration, electric potential, or both. 1 Channels permit movement of specific ions (or water) down their electrochemical gradient. 2 Transporters, which fall into three groups, facilitate movement of specific small molecules or ions. Uniporters transport a single type of molecule down its concentration gradient 2A. Cotransport proteins (symporters, 2B, and antiporters, 2C) catalyze the movement of one molecule against its concentration gradient (black circles), driven by movement of one or more ions down an electrochemical gradient (red circles). 3 Pumps use the energy released by ATP hydrolysis to power movement of specific ions or small molecules (red circles) against their electrochemical gradient. Differences in the mechanisms of transport by these three major classes of proteins account for their varying rates of solute movement.
Page 476
Channels transport water, specific ions, or hydrophilic small molecules across membranes down their concentration or electric potential gradients. Because this process requires transport proteins but not energy, it is sometimes referred to as passive transport or facilitated diffusion, but it is more properly called facilitated transport. Channels form a hydrophilic “tube” or passageway across the membrane through which multiple water molecules or ions move simultaneously, single file, at a very rapid rate. Some channels are open much of the time; they are referred to as nongated channels. Most ion channels, however, open only in response to specific chemical or electrical signals. These channels are referred to as gated channels because a protein “gate” alternatively blocks the channel or moves out of the way to open the channel (see Figure 11-2). Channels, like all transport proteins, are very selective for the type of molecule they transport.
Transporters (also called carriers) move a wide variety of ions and molecules across cellular membranes, but at a much slower rate than channels. Three types of transporters have been identified. Uniporters transport a single type of molecule down its concentration gradient. Glucose and amino acids cross the plasma membrane into most mammalian cells with the aid of uniporters. Collectively, channels and uniporters are sometimes called facilitated transporters, indicating movement down a concentration or electrochemical gradient.
In contrast, antiporters and symporters couple the movement of one type of ion or molecule against its concentration gradient with the movement of one or more different ions down its concentration gradient, in the same (symporter) or different (antiporter) directions. These proteins are often called cotransporters because of their ability to transport two or more different solutes simultaneously.
ATP-powered pumps (or simply pumps) are ATPases that use the energy of ATP hydrolysis to move ions or small molecules across a membrane against a chemical concentration gradient, an electric potential, or both. This process, referred to as active transport, is an example of coupled chemical reactions (see Chapter 2). In this case, transport of ions or small molecules “uphill” against an electrochemical gradient, which requires energy, is coupled to the hydrolysis of ATP, which releases energy. The overall reaction—ATP hydrolysis and the “uphill” movement of ions or small molecules—is energetically favorable.
Like cotransporters, pumps mediate coupled reactions in which an energetically unfavorable reaction (i.e., the uphill movement of one type of molecule or ion) is coupled to an energetically favorable reaction (i.e., the downhill movement of another). Note, however, that the nature of the energy-supplying reaction driving active transport by these two classes of proteins differs. ATP pumps use energy from hydrolysis of ATP, whereas cotransporters use the energy stored in an electrochemical gradient. The latter process is sometimes referred to as secondary active transport.
Conformational changes are essential to the function of all transport proteins. ATP-powered pumps and transporters undergo a cycle of conformational change exposing a binding site (or sites) to one side of the membrane in one conformation and to the other side in a second conformation. Because each such cycle results in the movement of only one substrate molecule (or, at most, a few), these proteins are characterized by relatively slow rates of transport, ranging from 100 to 104 ions or molecules per second (see Figure 11-2). Most ion channels shuttle between a closed state and an open state, but many ions can pass through an open channel without any further conformational change. For this reason, channels are characterized by very fast rates of transport, up to 108 ions per second.
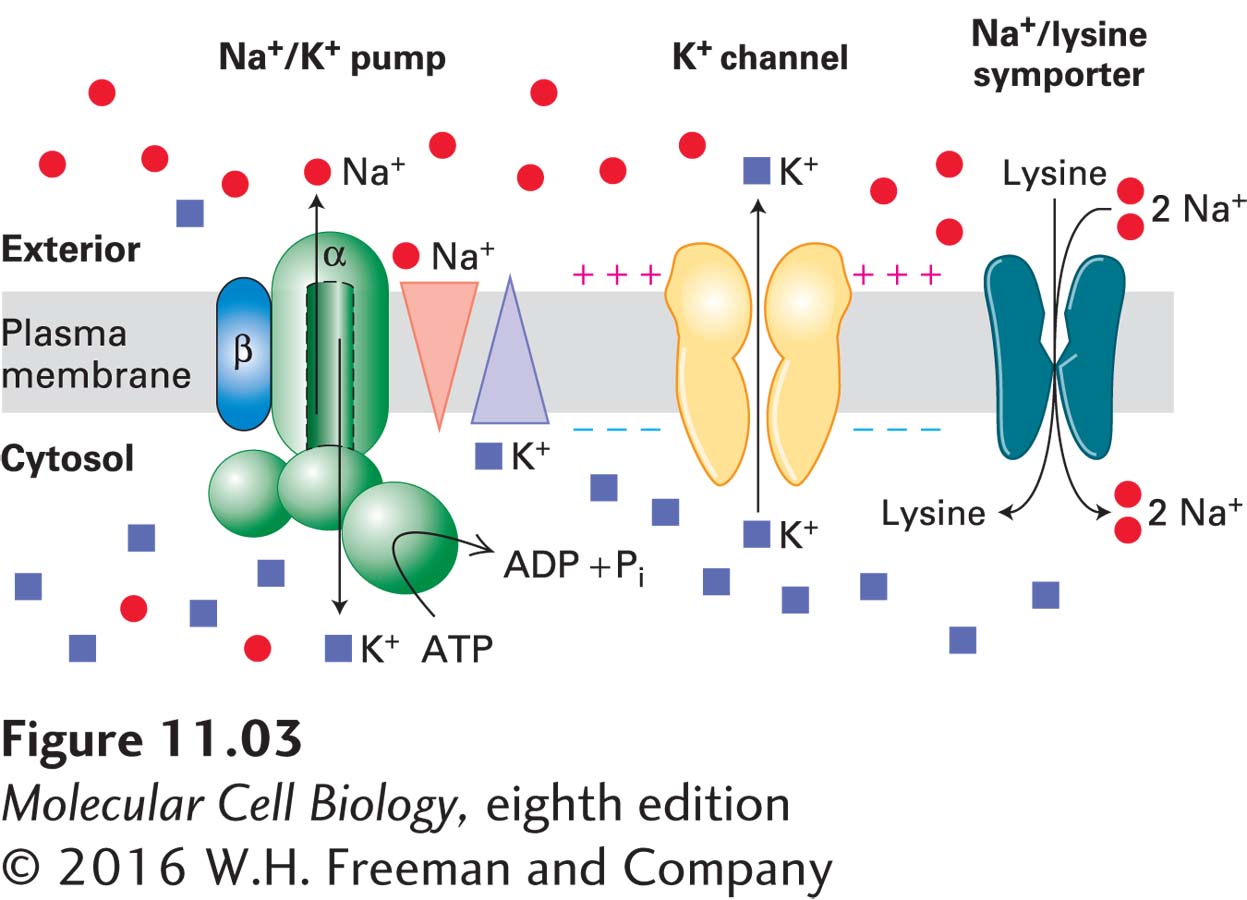
Frequently, several different types of transport proteins work in concert to achieve a physiological function. An example is seen in Figure 11-3, where an ATPase pumps Na+ out of the cell and K+ inward; this pump, which is found in virtually all metazoan cells, establishes the oppositely directed concentration gradients of Na+ and K+ ions across the plasma membrane (relatively high concentrations of K+ inside and Na+ outside cells) that are used to power the import of amino acids. The human genome encodes hundreds of different types of transport proteins that use the energy stored across the plasma membrane in the Na+ concentration gradient and its associated electric potential to transport a wide variety of molecules into cells against their concentration gradients.
FIGURE 11-3Multiple membrane transport proteins function together in the plasma membrane of metazoan cells. Gradients are indicated by triangles with the tip pointing toward lower concentration. The Na+/K+ ATPase in the plasma membrane uses energy released by ATP hydrolysis to pump Na+ (red circles) out of the cell and K+ (blue squares) inward; this creates a concentration gradient of Na+ that is greater outside the cell than inside, and one of K+ that is greater inside than outside. Movement of positively charged K+ ions out of the cell through membrane K+ channels creates an electric potential across the plasma membrane—the cytosolic face is negative with respect to the extracellular face. A Na+/lysine transporter, a typical sodium/amino acid cotransporter, moves two Na+ ions together with one lysine from the extracellular medium into the cell. “Uphill” movement of the amino acid is powered by “downhill” movement of Na+ ions, which in turn is powered both by the outside-greater-than-inside Na+ concentration gradient and by the negative charge on the inside of the plasma membrane, which attracts the positively charged Na+ ions. The ultimate source of the energy to power amino acid uptake comes from the ATP hydrolyzed by the Na+/K+ ATPase, since this pump creates both the Na+ ion concentration gradient and, via the K+ channels, the membrane potential, which together power the influx of Na+ ions.
Page 477
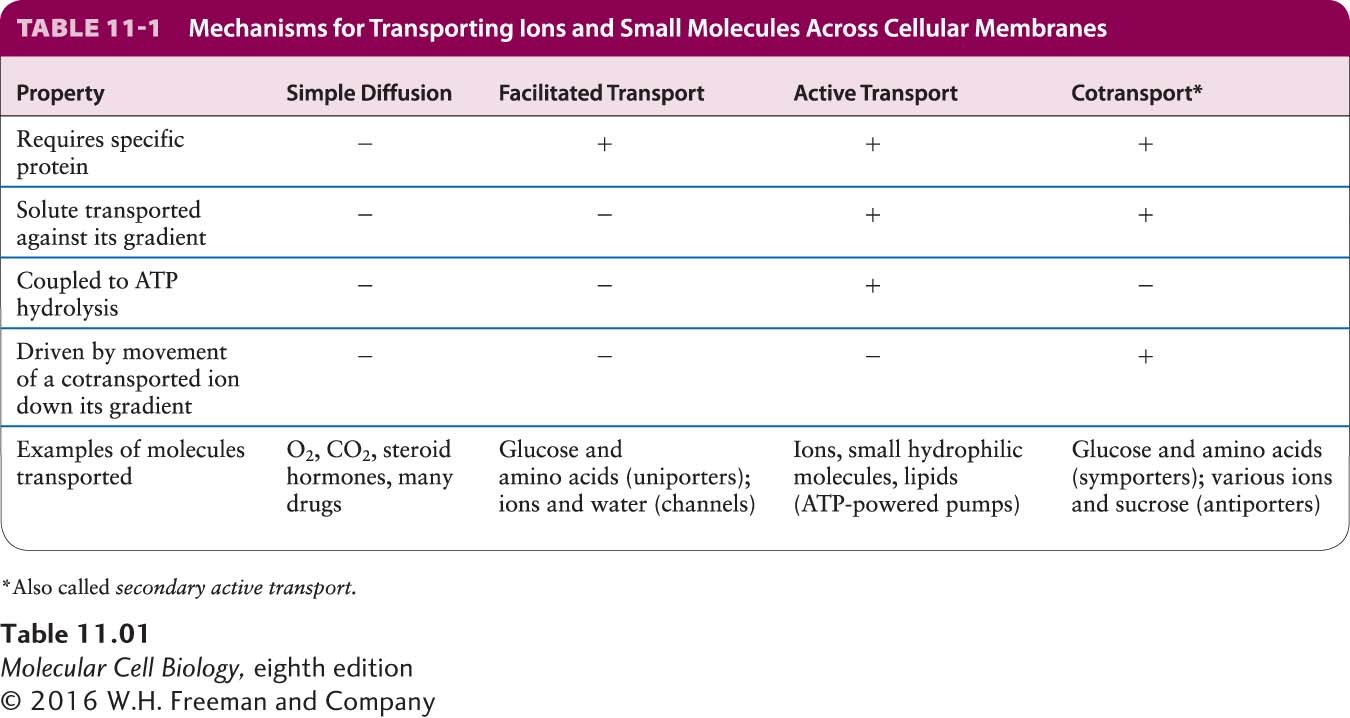
Table 11-1 summarizes the four mechanisms by which small molecules and ions are transported across cellular membranes. In the next section, we consider some of the simplest membrane transport proteins, those responsible for the transport of glucose and water.