Multipass Proteins Have Multiple Internal Topogenic Sequences
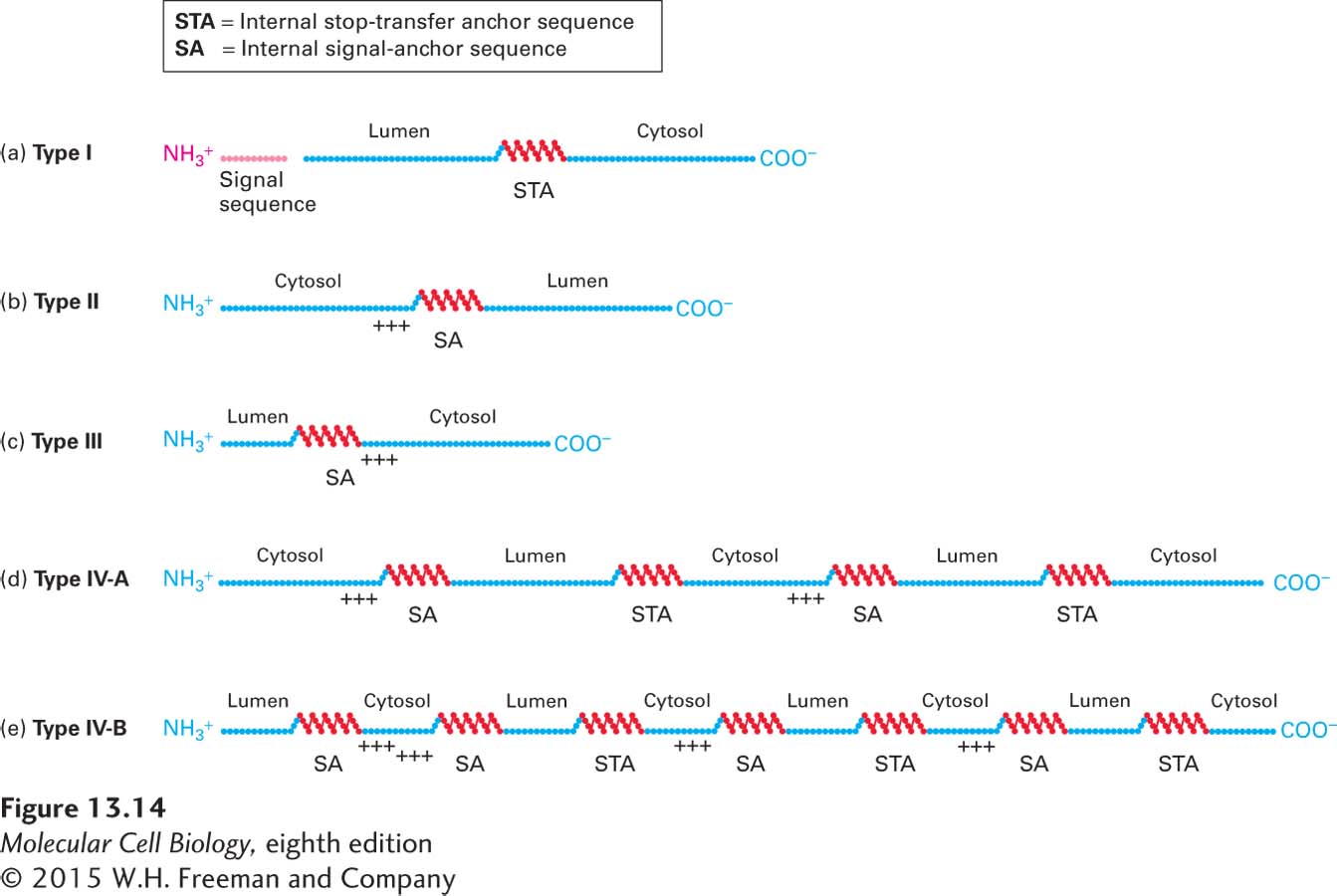
Figure 13-14 summarizes the arrangements of topogenic sequences in single-pass and multipass transmembrane proteins. In multipass (type IV) proteins, each of the membrane-spanning α helices acts as a topogenic sequence in the ways that we have already discussed: it can act to direct the protein to the ER, to anchor the protein in the ER membrane, or to stop transfer of the protein through the membrane. Multipass proteins fall into one of two types, depending on whether the N-terminus extends into the cytosol or the exoplasmic space (e.g., the ER lumen or cell exterior). This N-terminal topology is usually determined by the hydrophobic segment closest to the N-terminus and the charge of the sequences flanking it. If a type IV protein has an even number of transmembrane α helices, its N-terminus and C-terminus will be oriented toward the same side of the membrane (Figure 13-14d). Conversely, if a type IV protein has an odd number of α helices, its two ends will have opposite orientations (Figure 13-14e).
FIGURE 13-14Topogenic sequences determine the orientation of ER membrane proteins. Topogenic sequences are shown in red; soluble, hydrophilic sequences in blue. The internal topogenic sequences form transmembrane α helices that anchor proteins or segments of proteins in the membrane. (a) Type I proteins contain a cleaved signal sequence and a single internal stop-transfer anchor (STA) sequence. (b, c) Type II and type III proteins contain a single internal signal-anchor (SA) sequence. The difference in the orientation of these protein types depends largely on whether there is a high density of positively charged amino acids (+ + +) on the N-terminal side (type II) or on the C-terminal side of the SA sequence (type III). (d, e) Nearly all multipass proteins lack a cleavable signal sequence, as depicted in the examples shown here. Type IV-A proteins, whose N-terminus faces the cytosol, contain alternating type II SA sequences and STA sequences. Type IV-B proteins, whose N-terminus faces the lumen, begin with a type III SA sequence followed by alternating type II SA and STA sequences. Proteins of each type with different numbers of α helices (odd or even) are known.
Type IV Proteins with N-Terminus in the Cytosol Among the multipass proteins whose N-terminus extends into the cytosol are the various glucose transporters (GLUTs) and most ion-channel proteins, discussed in Chapter 11. In these proteins, the hydrophobic segment closest to the N-terminus initiates insertion of the nascent polypeptide chain into the ER membrane with the N-terminus oriented toward the cytosol; thus this α-helical segment functions like the internal signal-anchor sequence of a type II protein (see Figure 13-12a). As the nascent chain following the first α helix elongates, it moves through the translocon until the second hydrophobic α helix is formed. This helix prevents further extrusion of the nascent chain through the translocon; thus its function is similar to that of the stop-transfer anchor sequence in a type I protein (see Figure 13-11).
After synthesis of the first two transmembrane α helices, both ends of the nascent chain face the cytosol, and the loop between them extends into the ER lumen. The C-terminus of the nascent chain then continues to grow into the cytosol, as it does in synthesis of type I and type III proteins. The third transmembrane α helix acts as another type II signal-anchor sequence and the fourth as another stop-transfer anchor sequence (see Figure 13-14d). Apparently, once the first topogenic sequence of a multipass polypeptide initiates association with the translocon, the ribosome remains attached to the translocon, and topogenic sequences that subsequently emerge from the ribosome are threaded into the translocon without the need for the SRP and the SRP receptor. In a manner that is not well understood, as new hydrophobic topogenic sequences engage the translocon, the previously engaged sequences move laterally out of the translocon using the same mechanism as for type I, type II, and type III membrane proteins.
Page 598
Experiments that use recombinant DNA techniques to exchange hydrophobic α helices have provided insight into the functioning of the topogenic sequences in type IV-A multipass proteins. These experiments indicate that the order of the hydrophobic α helices relative to one another in the growing chain largely determines whether a given helix functions as a signal-anchor sequence or a stop-transfer anchor sequence. Other than its hydrophobicity, the specific amino acid sequence of a particular helix has little bearing on its function. Thus the first N-terminal α helix and the subsequent odd-numbered ones function as signal-anchor sequences, whereas the intervening even-numbered helices function as stop-transfer anchor sequences. This odd-even relationship among signal-anchor and stop-transfer anchor sequences is dictated by the fact that the transmembrane α helices assume alternating orientations as a multipass protein is woven back and forth across the membrane; signal-anchor sequences are oriented with their N-termini toward the cytoplasmic side of the bilayer, whereas stop-transfer anchor sequences have their N-termini oriented toward the exoplasmic side of the bilayer.
Type IV Proteins with N-Terminus in the Exoplasmic Space The large family of G protein–coupled receptors, all of which contain seven transmembrane α helices, constitute the most numerous type IV-B proteins, whose N-terminus extends into the exoplasmic space. In these proteins, the hydrophobic α helix closest to the N-terminus is often followed by a cluster of positively charged amino acids, like a type III signal-anchor sequence (see Figure 13-12b). As a result, the nascent polypeptide chain is inserted into the translocon with the N-terminus extending into the lumen (see Figure 13-14e). As the chain is elongated, it is inserted into the ER membrane by alternating type II signal-anchor sequences and stop-transfer sequences, as just described for type IV-A proteins.