Chaperones and Other ER Proteins Facilitate Folding and Assembly of Proteins
Although many denatured proteins can spontaneously refold into their native state in vitro, such refolding usually requires hours to reach completion. Yet proteins produced in the ER generally fold into their proper conformation within minutes after their synthesis. The rapid folding of these newly synthesized proteins in cells depends on the sequential action of several proteins present within the ER lumen. We have already seen how the molecular chaperone BiP can drive post-translational translocation in yeast by binding fully synthesized polypeptides as they enter the ER (see Figure 13-9). BiP can also bind transiently to nascent polypeptide chains as they enter the ER during cotranslational translocation. Bound BiP is thought to prevent segments of a nascent chain from misfolding or forming aggregates, thereby promoting folding of the entire polypeptide into the proper conformation. PDI also contributes to proper folding because the correct three-dimensional conformation is stabilized by disulfide bonds in many proteins.
Page 605
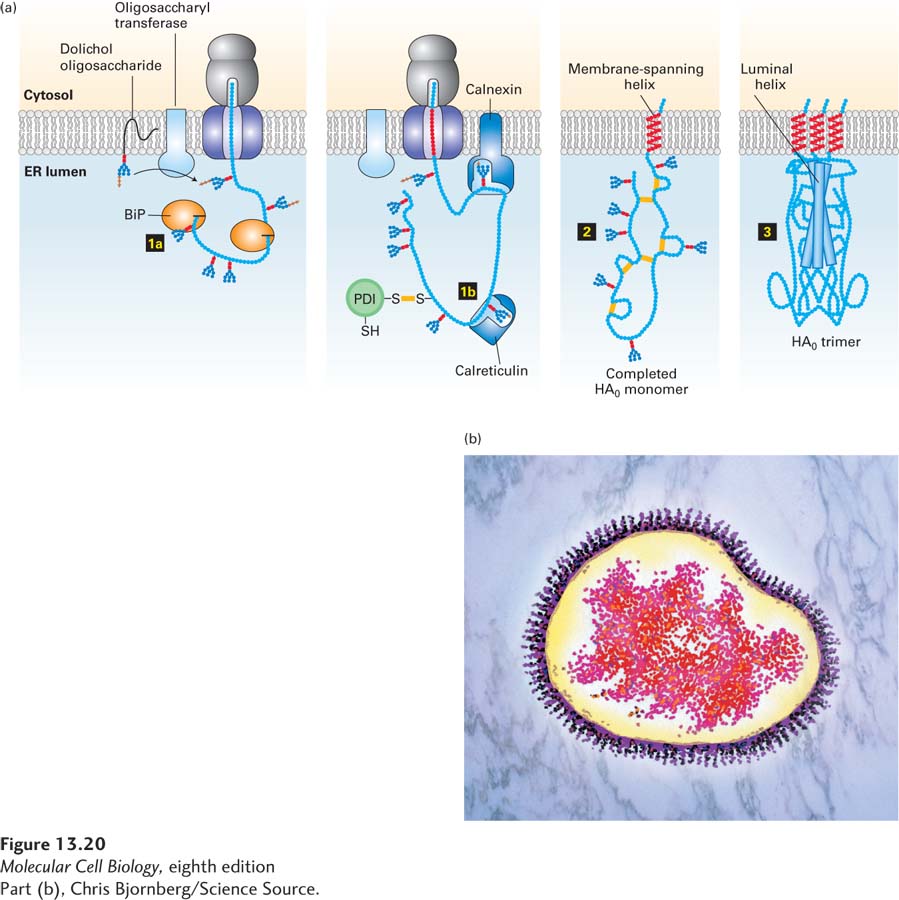
As illustrated in Figure 13-20, two other ER proteins, the homologous lectins (carbohydrate-binding proteins) calnexin and calreticulin, bind selectively to certain N-linked oligosaccharides on growing polypeptide chains. The ligand for these two proteins, which resembles the N-linked oligosaccharide precursor but has only a single glucose residue [Glc1Man9(GlcNAc)2], is generated by a specific glucosyltransferase in the ER lumen (see Figure 13-18, step 3a). This enzyme acts only on polypeptide chains that are unfolded or misfolded, and in this respect, the glucosyltransferase acts as one of the primary surveillance mechanisms to ensure quality control of protein folding in the ER. Unfolded proteins often expose hydrophobic segments that in a properly folded state are buried in the hydrophobic core of the protein. The glucosyltransferase probably recognizes unfolded proteins by binding to these exposed hydrophobic segments. Binding of calnexin and calreticulin to unfolded nascent chains marked with glucosylated N-linked oligosaccharides prevents aggregation of adjacent segments of a protein as it is being made on the ER. Thus calnexin and calreticulin, like BiP, help prevent premature, incorrect folding of segments of a newly made protein.
[Part (b), Chris Bjornberg/Science Source.]
FIGURE 13-20Hemagglutinin folding and assembly. (a) Mechanism of (HA0) trimer assembly. Transient binding of the chaperone BiP (step 1a) to the nascent polypeptide chain and of two lectins, calnexin and calreticulin, to certain oligosaccharide chains (step 1b) promotes proper folding of adjacent segments of HA0. A total of seven N-linked oligosaccharide chains are added to the luminal portion of the nascent chain during cotranslational translocation, and PDI catalyzes the formation of six disulfide bonds per monomer. Completed HA0 monomers are anchored in the membrane by a single membrane-spanning α helix with the N-terminus in the lumen (step 2). Interaction of three HA0 chains with one another, initially via their transmembrane α helices, apparently triggers formation of a long stem containing one α helix from the luminal part of each HA0 polypeptide. Finally, interactions occur among the three globular heads, generating a stable HA0 trimer (step 3). (b) Electron micrograph (false color) of a complete influenza virion showing trimers of HA protein protruding as spikes from the surface of the viral membrane. See U. Tatu et al., 1995, EMBO J.14:1340, and D. Hebert et al., 1997, J. Cell Biol.139:613.
[Part (b), Chris Bjornberg/Science Source.]
Page 606
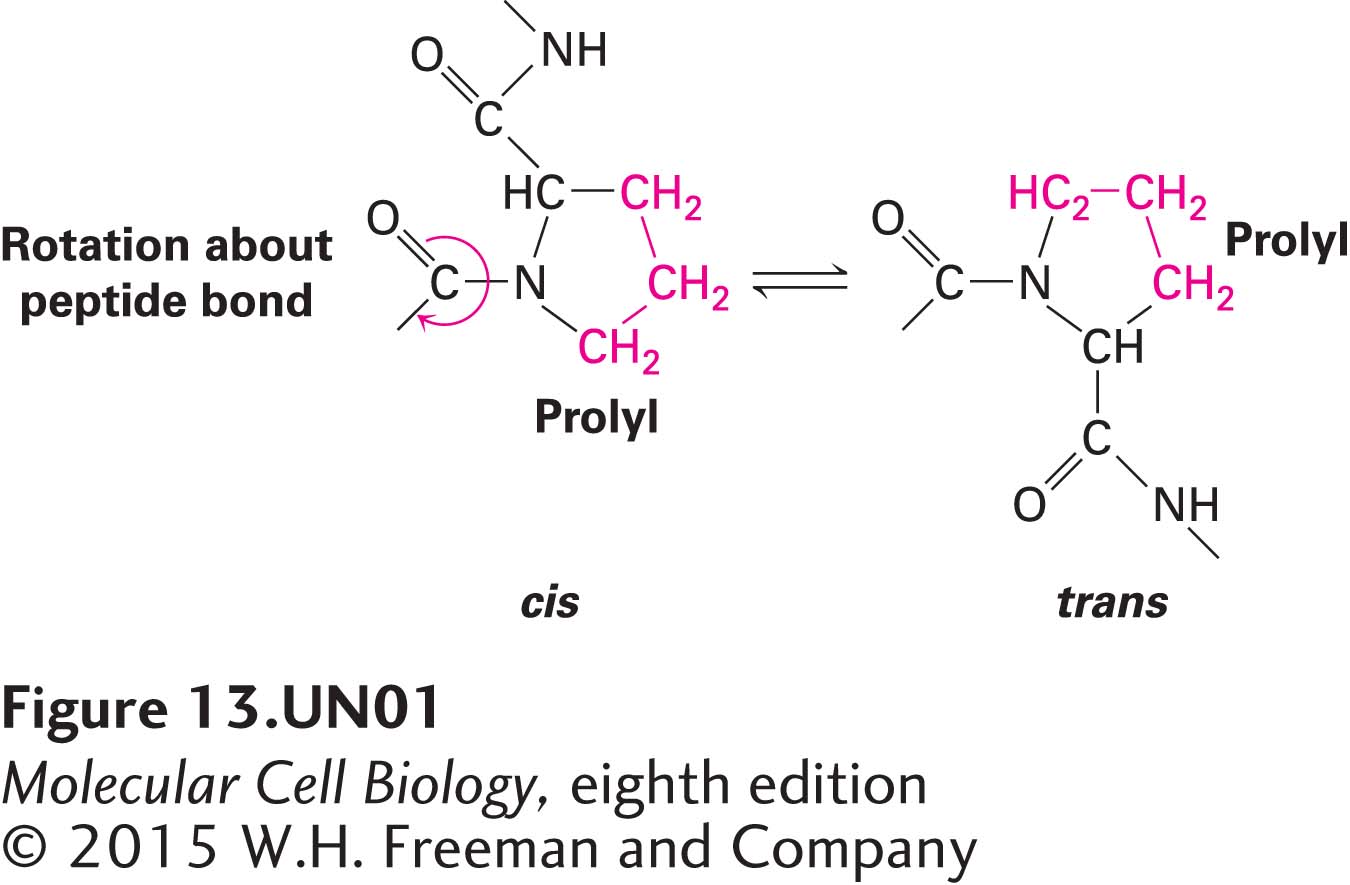
Other important protein-folding catalysts in the ER lumen are peptidyl-prolyl isomerases, a family of enzymes that accelerate the rotation about peptidyl-prolyl bonds at proline residues in unfolded segments of a polypeptide:
Such isomerizations are sometimes the rate-limiting step in the folding of protein domains. Many peptidyl-prolyl isomerases can catalyze the rotation of exposed peptidyl-prolyl bonds indiscriminately in numerous proteins, but some have very specific protein substrates.
Many important soluble secretory and membrane proteins synthesized on the ER are built of two or more polypeptide subunits. In all cases, the assembly of the subunits constituting these multisubunit (multimeric) proteins occurs in the ER. The immunoglobulins, which contain two heavy (H) and two light (L) chains, all linked by intrachain disulfide bonds, are assembled in this way. Hemagglutinin (HA) is another multimeric protein that provides a good illustration of folding and subunit assembly (see Figure 13-20). This trimeric protein forms the spikes that protrude from the surface of an influenza virus particle. The HA trimer is formed within the ER of an infected host cell from three copies of a precursor protein termed HA0, which has a single membrane-spanning α helix. In the Golgi complex, each of the three HA0 proteins is cleaved to form two polypeptides, HA1 and HA2; thus each HA molecule that eventually resides on the viral surface contains three copies of HA1 and three of HA2 (see Figure 3-11). The trimer is stabilized by interactions between the large exoplasmic domains of the constituent polypeptides, which extend into the ER lumen; after HA is transported to the cell surface, these domains extend into the extracellular space. Interactions between the smaller cytosolic and membrane-spanning portions of the HA subunits also help stabilize the trimeric protein. Studies have shown that it takes just 10 minutes for the HA0 polypeptides to fold and assemble into their proper trimeric conformation.